

Mechanism of inhibition of enveloped virus membrane fusion by the antiviral drug arbidol
- PMID: 21283579
- PMCID: PMC3026800
- DOI: 10.1371/journal.pone.0015874
Mechanism of inhibition of enveloped virus membrane fusion by the antiviral drug arbidol
Abstract
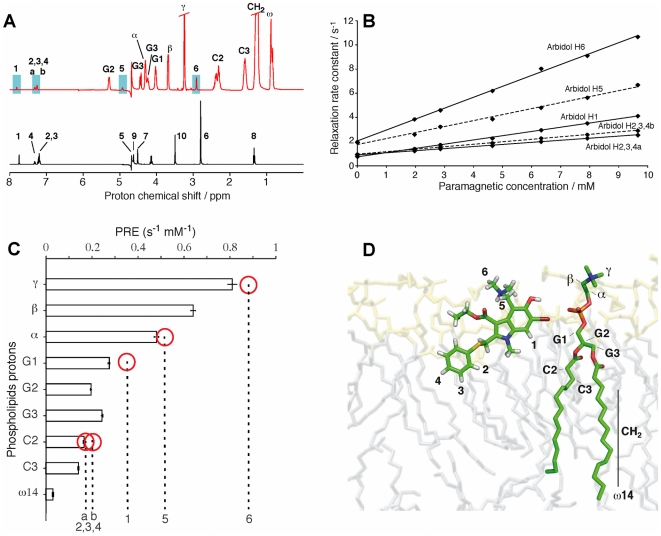
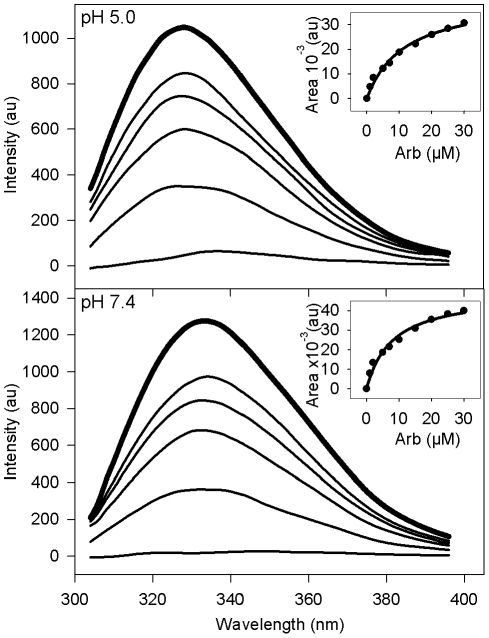
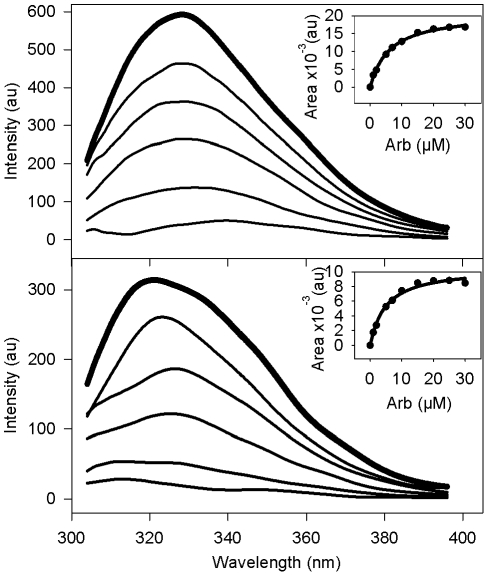
The broad-spectrum antiviral arbidol (Arb) inhibits cell entry of enveloped viruses by blocking viral fusion with host cell membrane. To better understand Arb mechanism of action, we investigated its interactions with phospholipids and membrane peptides. We demonstrate that Arb associates with phospholipids in the micromolar range. NMR reveals that Arb interacts with the polar head-group of phospholipid at the membrane interface. Fluorescence studies of interactions between Arb and either tryptophan derivatives or membrane peptides reconstituted into liposomes show that Arb interacts with tryptophan in the micromolar range. Interestingly, apparent binding affinities between lipids and tryptophan residues are comparable with those of Arb IC50 of the hepatitis C virus (HCV) membrane fusion. Since tryptophan residues of membrane proteins are known to bind preferentially at the membrane interface, these data suggest that Arb could increase the strength of virus glycoprotein's interactions with the membrane, due to a dual binding mode involving aromatic residues and phospholipids. The resulting complexation would inhibit the expected viral glycoprotein conformational changes required during the fusion process. Our findings pave the way towards the design of new drugs exhibiting Arb-like interfacial membrane binding properties to inhibit early steps of virus entry, i.e., attractive targets to combat viral infection.
Conflict of interest statement
Figures
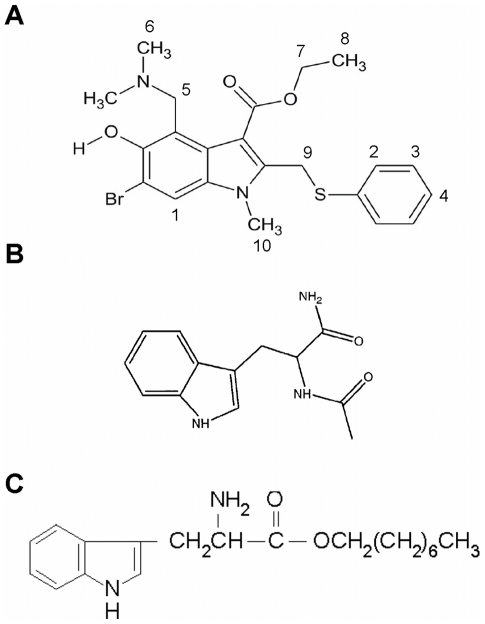
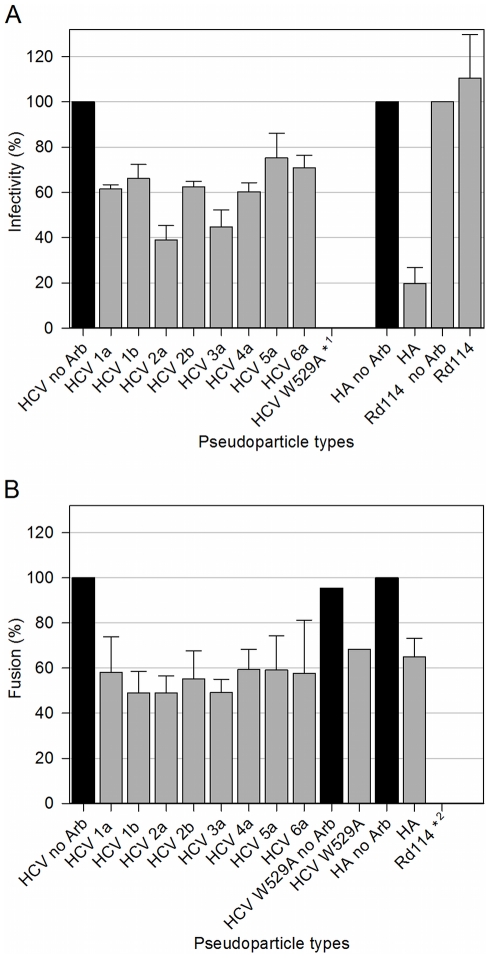
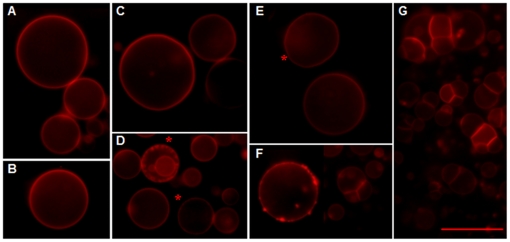
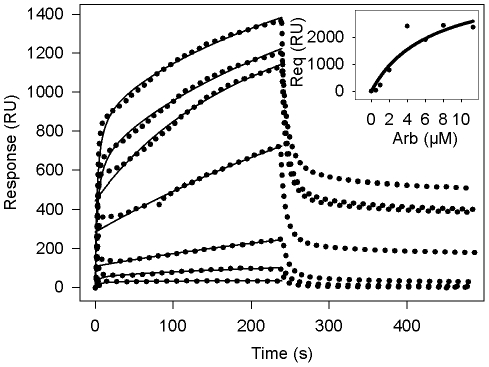
References
-
- Ahmed-Belkacem A, Ahnou N, Barbotte L, Wychowski C, Pallier C, et al. Silibinin and related compounds are direct inhibitors of hepatitis C virus RNA-dependent RNA polymerase. Gastroenterology. 2010;138:1112–1122. - PubMed
-
- Chai H, Zhao Y, Zhao C, Gong P. Synthesis and in vitro anti-hepatitis B virus activities of some ethyl 6-bromo-5-hydroxy-1H-indole-3-carboxylates. Bioorg Med Chem. 2006;14:911–917. - PubMed
-
- Brooks MJ, Sasadeusz JJ, Tannock GA. Antiviral chemotherapeutic agents against respiratory viruses: where are we now and what's in the pipeline? Curr Opin Pulm Med. 2004;10:197–203. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
