

The Hsc/Hsp70 co-chaperone network controls antigen aggregation and presentation during maturation of professional antigen presenting cells
- PMID: 21283720
- PMCID: PMC3024426
- DOI: 10.1371/journal.pone.0016398
The Hsc/Hsp70 co-chaperone network controls antigen aggregation and presentation during maturation of professional antigen presenting cells
Abstract
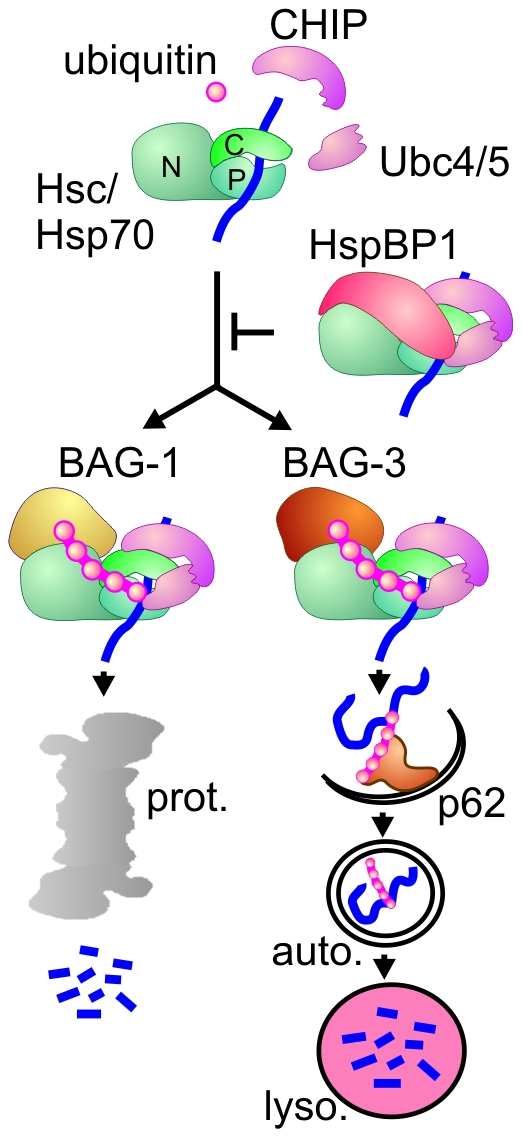
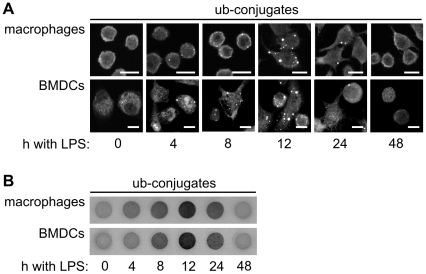
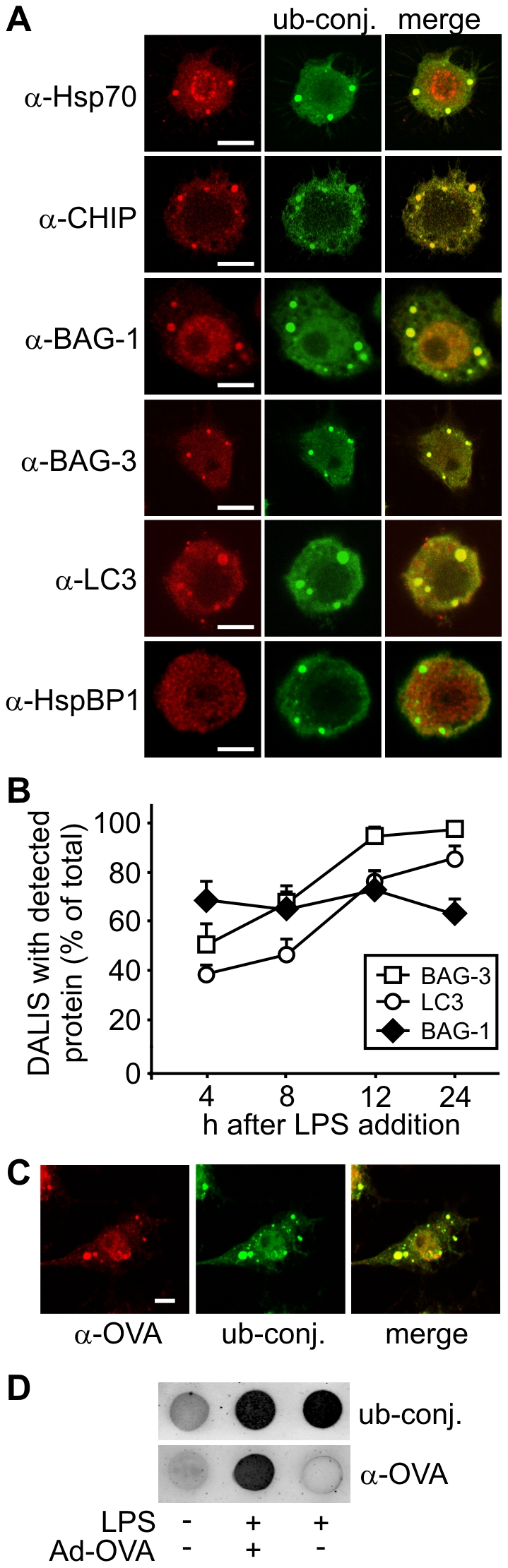
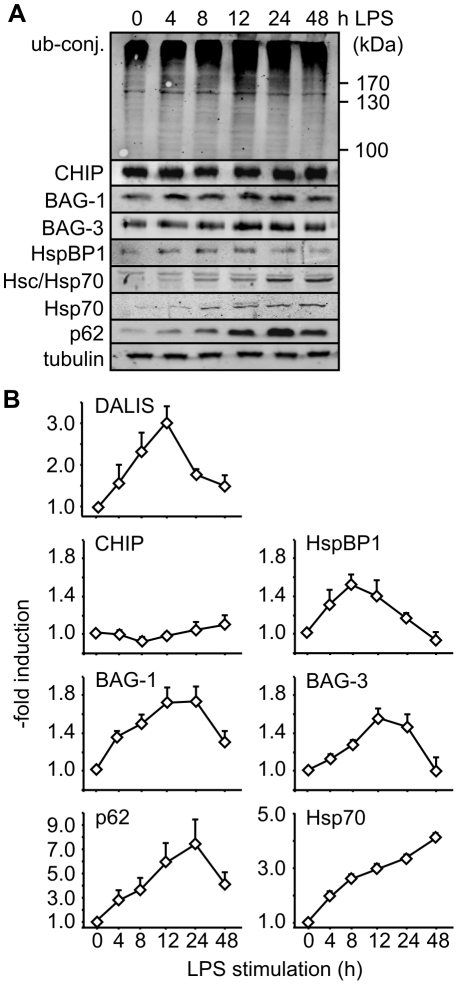
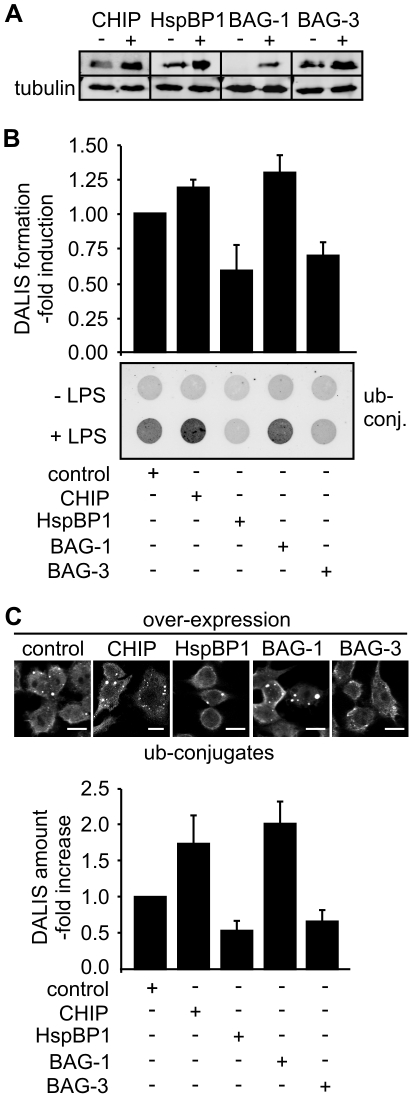
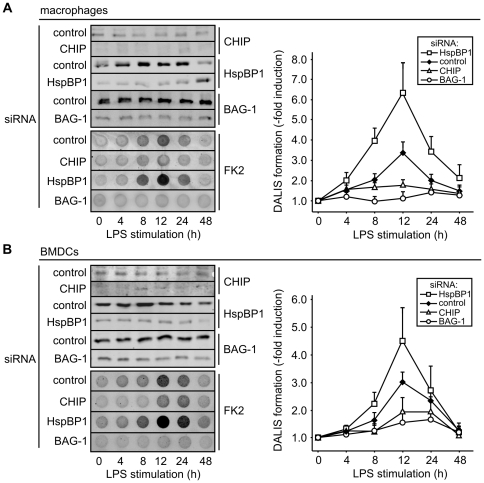
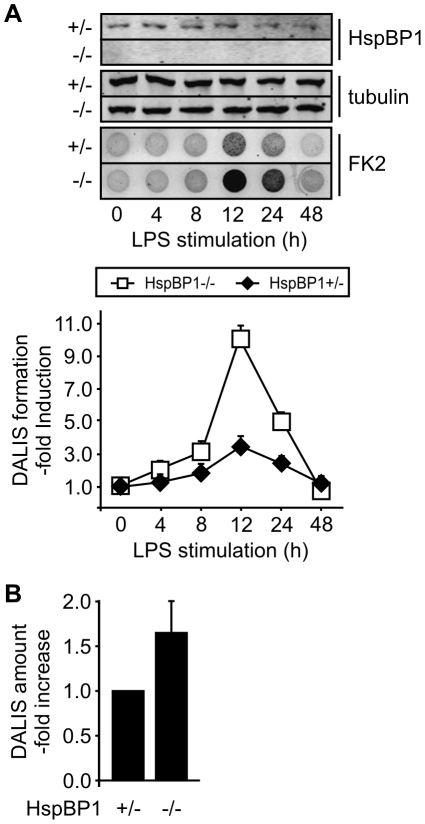
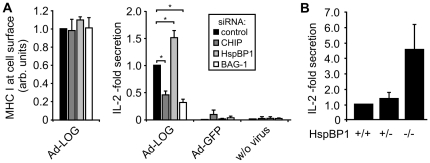
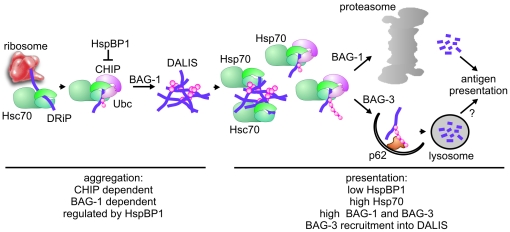
The maturation of mouse macrophages and dendritic cells involves the transient deposition of ubiquitylated proteins in the form of dendritic cell aggresome-like induced structures (DALIS). Transient DALIS formation was used here as a paradigm to study how mammalian cells influence the formation and disassembly of protein aggregates through alterations of their proteostasis machinery. Co-chaperones that modulate the interplay of Hsc70 and Hsp70 with the ubiquitin-proteasome system (UPS) and the autophagosome-lysosome pathway emerged as key regulators of this process. The chaperone-associated ubiquitin ligase CHIP and the ubiquitin-domain protein BAG-1 are essential for DALIS formation in mouse macrophages and bone-marrow derived dendritic cells (BMDCs). CHIP also cooperates with BAG-3 and the autophagic ubiquitin adaptor p62 in the clearance of DALIS through chaperone-assisted selective autophagy (CASA). On the other hand, the co-chaperone HspBP1 inhibits the activity of CHIP and thereby attenuates antigen sequestration. Through a modulation of DALIS formation CHIP, BAG-1 and HspBP1 alter MHC class I mediated antigen presentation in mouse BMDCs. Our data show that the Hsc/Hsp70 co-chaperone network controls transient protein aggregation during maturation of professional antigen presenting cells and in this way regulates the immune response. Similar mechanisms may modulate the formation of aggresomes and aggresome-like induced structures (ALIS) in other mammalian cell types.
Conflict of interest statement
Figures
References
-
- Balch WE, Morimoto RI, Dillin A, Kelly JW. Adapting proteostasis for disease intervention. Science. 2008;319:916–919. - PubMed
-
- Brodsky JL, Chiosis G. Hsp70 molecular chaperones: emerging roles in human disease and identification of small molecule modulators. Curr Top Med Chem. 2006;6:1215–1225. - PubMed
-
- Hartl FU, Hayer-Hartl M. Converging concepts of protein folding in vitro and in vivo. Nat Struct Mol Biol. 2009;16:574–581. - PubMed
-
- Kettern N, Dreiseidler M, Tawo R, Höhfeld J. Chaperone-assisted degradation: multiple paths to destruction. Biol Chem. 2010;391:481–489. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
