

The gene regulatory cascade linking proneural specification with differentiation in Drosophila sensory neurons
- PMID: 21283833
- PMCID: PMC3023811
- DOI: 10.1371/journal.pbio.1000568
The gene regulatory cascade linking proneural specification with differentiation in Drosophila sensory neurons
Abstract
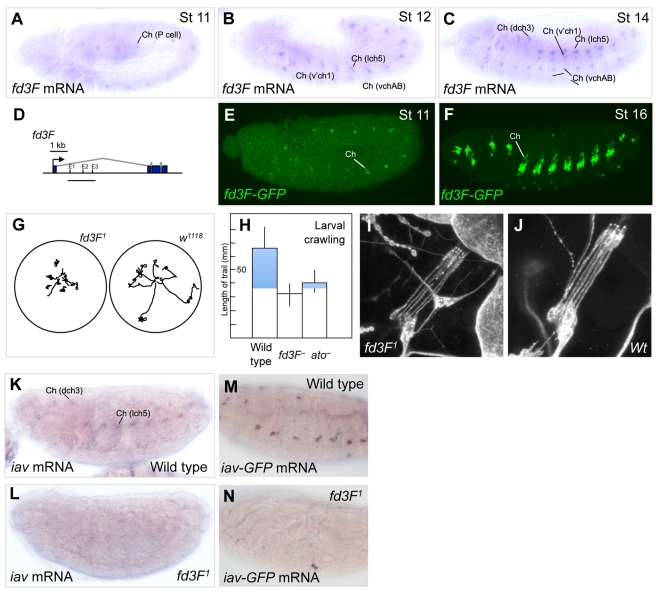
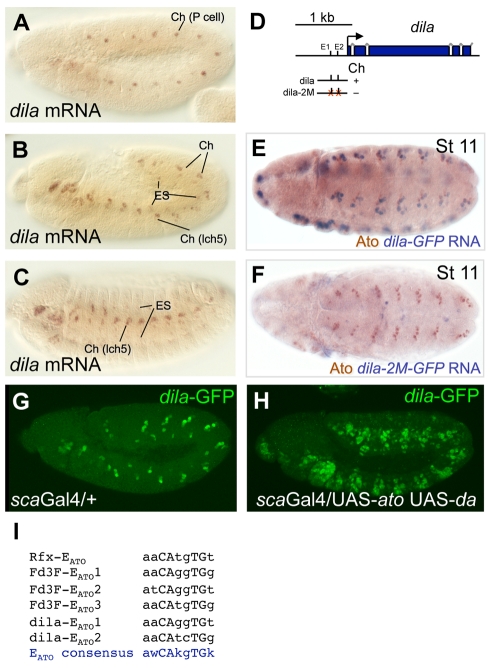
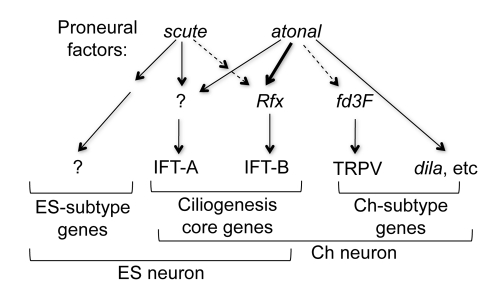
In neurogenesis, neural cell fate specification is generally triggered by proneural transcription factors. Whilst the role of proneural factors in fate specification is well studied, the link between neural specification and the cellular pathways that ultimately must be activated to construct specialised neurons is usually obscure. High-resolution temporal profiling of gene expression reveals the events downstream of atonal proneural gene function during the development of Drosophila chordotonal (mechanosensory) neurons. Among other findings, this reveals the onset of expression of genes required for construction of the ciliary dendrite, a key specialisation of mechanosensory neurons. We determine that atonal activates this cellular differentiation pathway in several ways. Firstly, atonal directly regulates Rfx, a well-known highly conserved ciliogenesis transcriptional regulator. Unexpectedly, differences in Rfx regulation by proneural factors may underlie variations in ciliary dendrite specialisation in different sensory neuronal lineages. In contrast, fd3F encodes a novel forkhead family transcription factor that is exclusively expressed in differentiating chordotonal neurons. fd3F regulates genes required for specialized aspects of chordotonal dendrite physiology. In addition to these intermediate transcriptional regulators, we show that atonal directly regulates a novel gene, dilatory, that is directly associated with ciliogenesis during neuronal differentiation. Our analysis demonstrates how early cell fate specification factors can regulate structural and physiological differentiation of neuronal cell types. It also suggests a model for how subtype differentiation in different neuronal lineages may be regulated by different proneural factors. In addition, it provides a paradigm for how transcriptional regulation may modulate the ciliogenesis pathway to give rise to structurally and functionally specialised ciliary dendrites.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
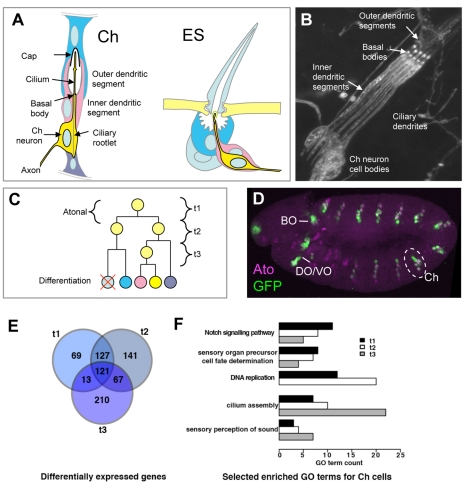
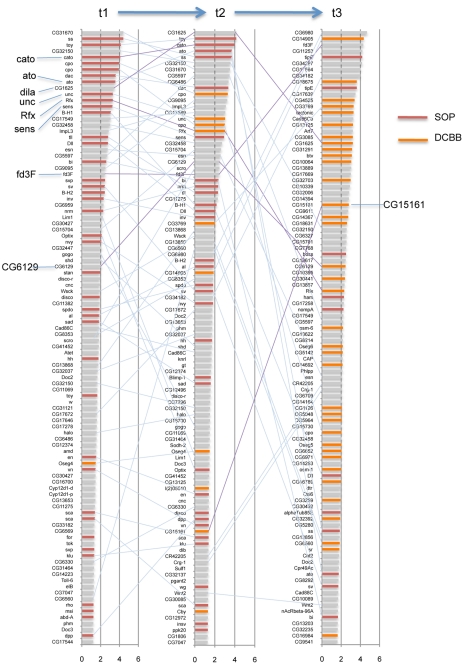
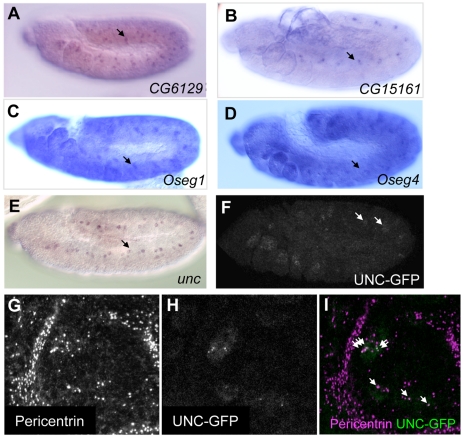
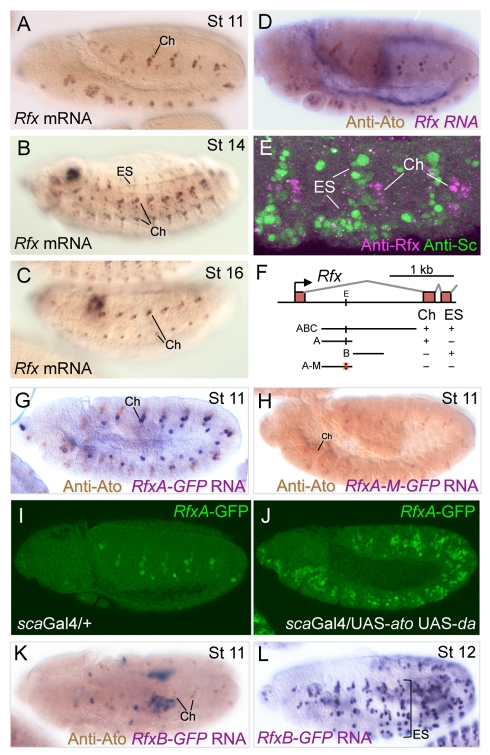
References
-
- Bertrand N, Castro D. S, Guillemot F. Proneural genes and the specification of neural cell types. Nature Rev Neuro. 2002;3:517–530. - PubMed
-
- Bermingham N. A, Hassan B. A, Price S. D, Vollrath M. A, Ben-Arie N, et al. Math1, an essential gene for the generation of inner ear hair cells. Science. 1999;284:1837–1841. - PubMed
-
- Ben-Arie N, Hassan B. A, Bermingham N. A, Malicki D. M, Armstrong D, et al. Functional conservation of atonal and Math1 in the CNS and PNS. Development. 2000;127:1039–1048. - PubMed
-
- Jarman A. P, Sun Y, Jan L. Y, Jan Y. N. Role of the proneural gene, atonal, in formation of Drosophila chordotonal organs and photoreceptors. Development. 1995;121:2019–2030. - PubMed
-
- Singson A, Leviten M. W, Bang A. G, Hua X. H, Posakony J. W. Direct downstream targets of proneural activators in the imaginal disc include genes involved in lateral inhibitory signaling. Genes and Development. 1994;8:2058–2071. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
