

The chromatin remodeling factor Bap55 functions through the TIP60 complex to regulate olfactory projection neuron dendrite targeting
- PMID: 21284845
- PMCID: PMC3038883
- DOI: 10.1186/1749-8104-6-5
The chromatin remodeling factor Bap55 functions through the TIP60 complex to regulate olfactory projection neuron dendrite targeting
Abstract
Background: The Drosophila olfactory system exhibits very precise and stereotyped wiring that is specified predominantly by genetic programming. Dendrites of olfactory projection neurons (PNs) pattern the developing antennal lobe before olfactory receptor neuron axon arrival, indicating an intrinsic wiring mechanism for PN dendrites. These wiring decisions are likely determined through a transcriptional program.
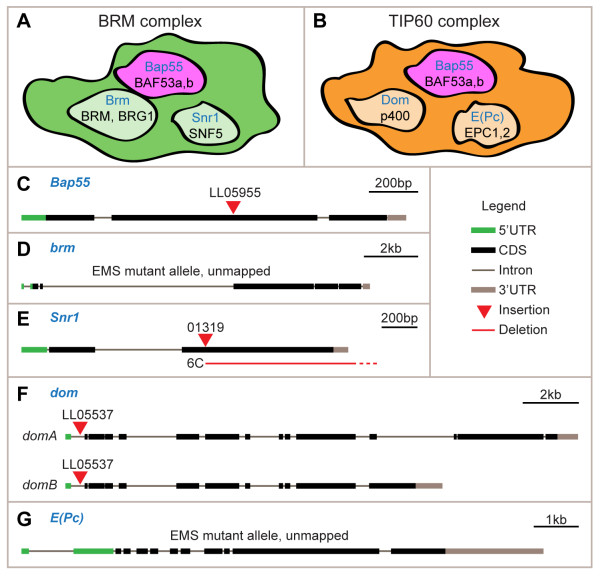
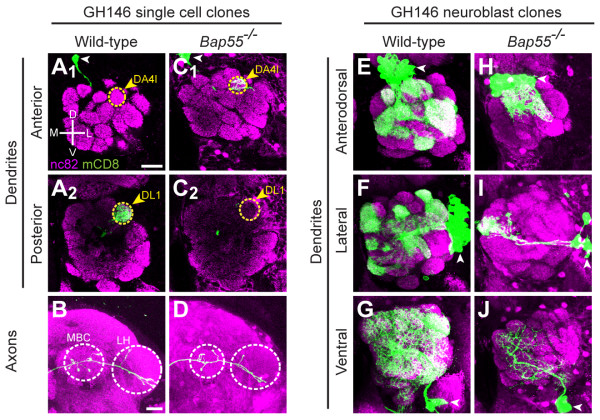
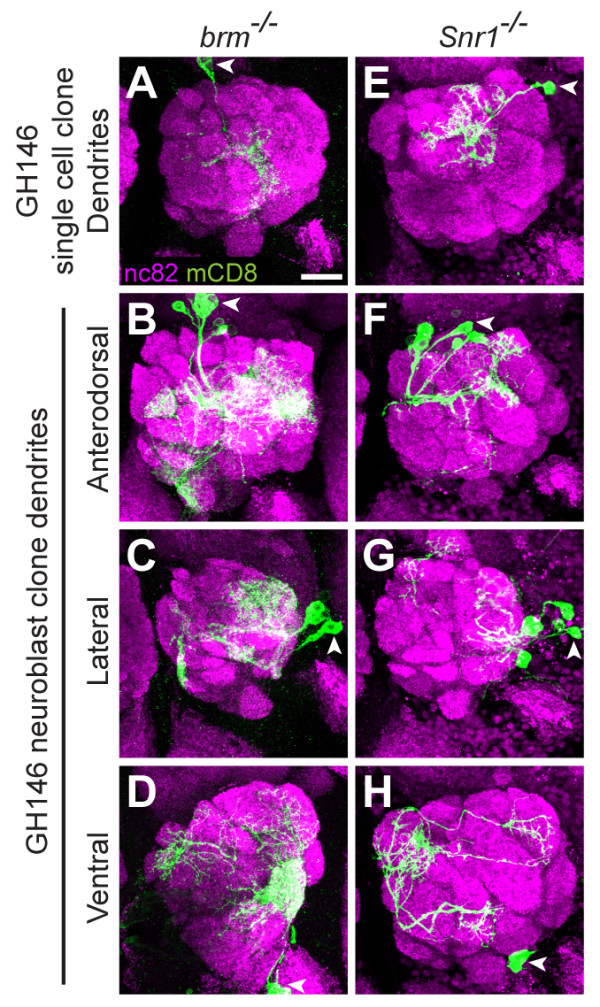
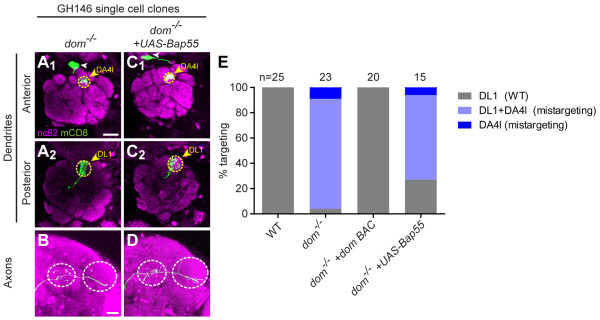
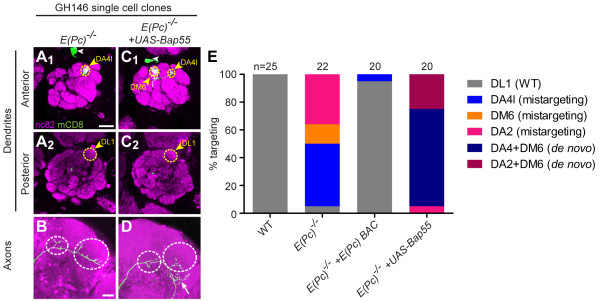
Results: We find that loss of Brahma associated protein 55 kD (Bap55) results in a highly specific PN mistargeting phenotype. In Bap55 mutants, PNs that normally target to the DL1 glomerulus mistarget to the DA4l glomerulus with 100% penetrance. Loss of Bap55 also causes derepression of a GAL4 whose expression is normally restricted to a small subset of PNs. Bap55 is a member of both the Brahma (BRM) and the Tat interactive protein 60 kD (TIP60) ATP-dependent chromatin remodeling complexes. The Bap55 mutant phenotype is partially recapitulated by Domino and Enhancer of Polycomb mutants, members of the TIP60 complex. However, distinct phenotypes are seen in Brahma and Snf5-related 1 mutants, members of the BRM complex. The Bap55 mutant phenotype can be rescued by postmitotic expression of Bap55, or its human homologs BAF53a and BAF53b.
Conclusions: Our results suggest that Bap55 functions through the TIP60 chromatin remodeling complex to regulate dendrite wiring specificity in PNs. The specificity of the mutant phenotypes suggests a position for the TIP60 complex at the top of a regulatory hierarchy that orchestrates dendrite targeting decisions.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
