

Distinct roles of deiodinases on the phenotype of Mct8 defect: a comparison of eight different mouse genotypes
- PMID: 21285310
- PMCID: PMC3040057
- DOI: 10.1210/en.2010-0900
Distinct roles of deiodinases on the phenotype of Mct8 defect: a comparison of eight different mouse genotypes
Abstract
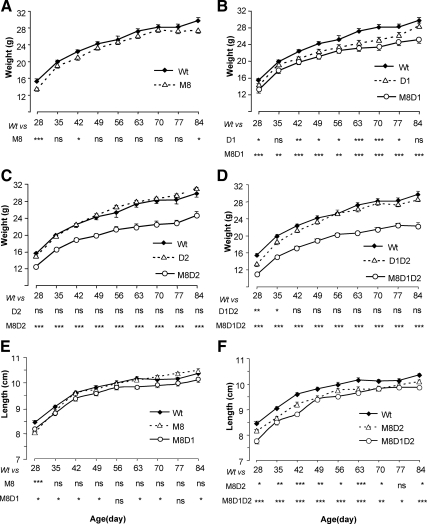
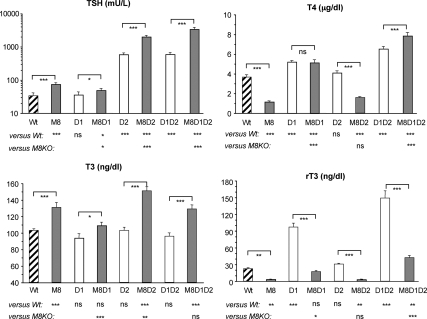
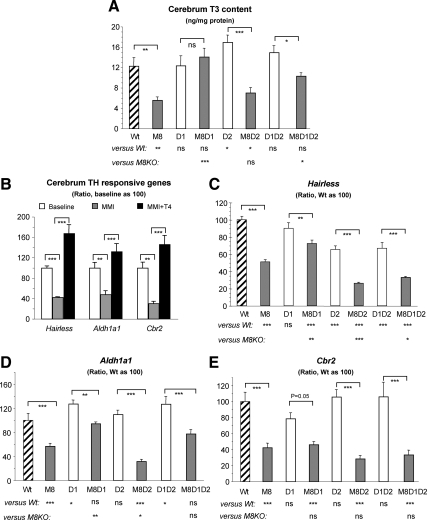
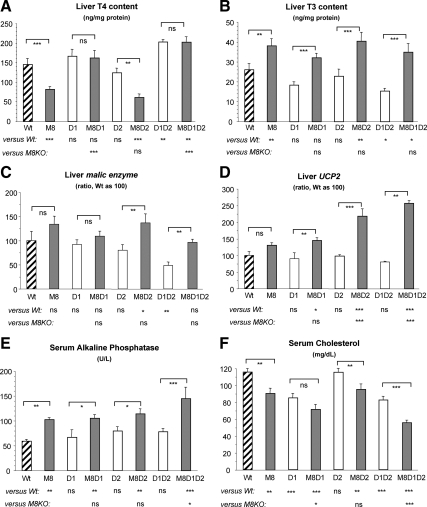
Mice deficient in the thyroid hormone (TH) transporter Mct8 (Mct8KO) have increased 5'-deiodination and impaired TH secretion and excretion. These and other unknown mechanisms result in the low-serum T(4), high T(3), and low rT(3) levels characteristic of Mct8 defects. We investigated to what extent each of the 5'-deiodinases (D1, D2) contributes to the serum TH abnormalities of the Mct8KO by generating mice with all combinations of Mct8 and D1 and/or D2 deficiencies and comparing the resulting eight genotypes. Adding D1 deficiency to that of Mct8 corrected the serum TH abnormalities of Mct8KO mice, normalized brain T(3) content, and reduced the impaired expression of TH-responsive genes. In contrast, Mct8D2KO mice maintained the serum TH abnormalities of Mct8KO mice. However, the serum TSH level increased 27-fold, suggesting a severely impaired hypothalamo-pituitary-thyroid axis. The brain of Mct8D2KO manifested a pattern of more severe impairment of TH action than Mct8KO alone. In triple Mct8D1D2KO mice, the markedly increased serum TH levels produced milder brain defect than that of Mct8D2KO at the expense of more severe liver thyrotoxicosis. Additionally, we observed that mice deficient in D2 had an unexplained marked reduction in the thyroid growth response to TSH. Our studies on these eight genotypes provide a unique insight into the complex interplay of the deiodinases in the Mct8 defect and suggest that D1 contributes to the increased serum T(3) in Mct8 deficiency, whereas D2 mainly functions locally, converting T(4) to T(3) to compensate for distinct cellular TH depletion in Mct8KO mice.
Figures
References
-
- Kopp P, Solis-Sainz JC. 2009. Thyroid hormone synthesis. In: Wondisford FE, Radovick S. eds. Clinical management of thyroid disease. Philadelphia: Saunders/Elsevier; 19–41
-
- Jansen J, Friesema EC, Milici C, Visser TJ. 2005. Thyroid hormone transporters in health and disease. Thyroid 15:757–768 - PubMed
-
- Bianco AC, Salvatore D, Gereben B, Berry MJ, Larsen PR. 2002. Biochemistry, cellular and molecular biology, and physiological roles of the iodothyronine selenodeiodinases. Endocr Rev 23:38–89 - PubMed
-
- Friesema EC, Ganguly S, Abdalla A, Manning Fox JE, Halestrap AP, Visser TJ. 2003. Identification of monocarboxylate transporter 8 as a specific thyroid hormone transporter. J Biol Chem 278:40128–40135 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
