

The curious case of NG2 cells: transient trend or game changer?
- PMID: 21288204
- PMCID: PMC3052864
- DOI: 10.1042/AN20110001
The curious case of NG2 cells: transient trend or game changer?
Abstract
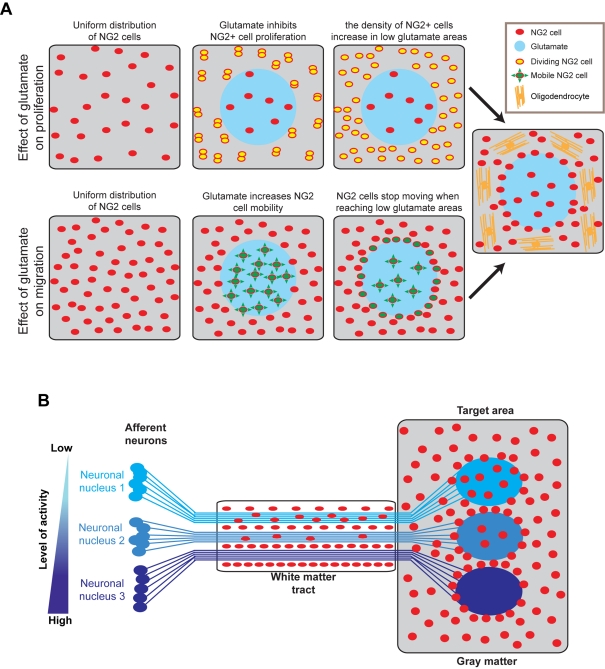
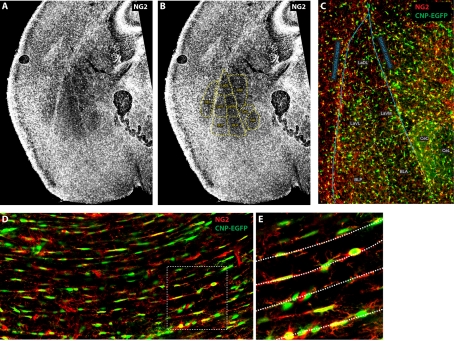
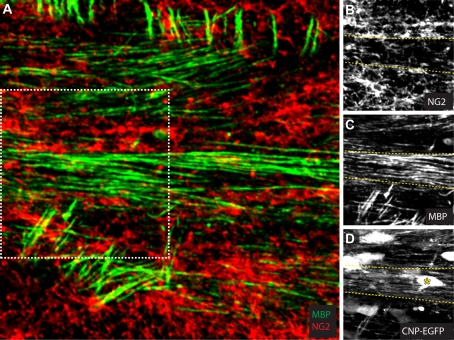
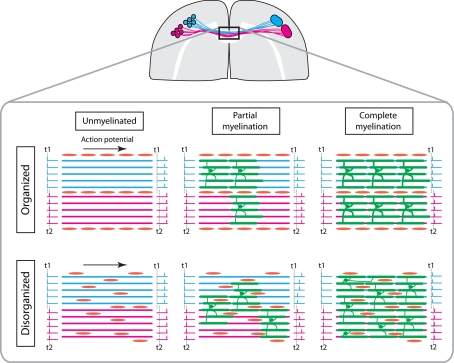
It has been 10 years since the seminal work of Dwight Bergles and collaborators demonstrated that NG2 (nerve/glial antigen 2)-expressing oligodendrocyte progenitor cells (NG2 cells) receive functional glutamatergic synapses from neurons (Bergles et al., 2000), contradicting the old dogma that only neurons possess the complex and specialized molecular machinery necessary to receive synapses. While this surprising discovery may have been initially shunned as a novelty item of undefined functional significance, the study of neuron-to-NG2 cell neurotransmission has since become a very active and exciting field of research. Many laboratories have now confirmed and extended the initial discovery, showing for example that NG2 cells can also receive inhibitory GABAergic synapses (Lin and Bergles, 2004) or that neuron-to-NG2 cell synaptic transmission is a rather ubiquitous phenomenon that has been observed in all brain areas explored so far, including white matter tracts (Kukley et al., 2007; Ziskin et al., 2007; Etxeberria et al., 2010). Thus, while still being in its infancy, this field of research has already brought many surprising and interesting discoveries, and has become part of a continuously growing effort in neuroscience to re-evaluate the long underestimated role of glial cells in brain function (Barres, 2008). However, this area of research is now reaching an important milestone and its long-term significance will be defined by its ability to uncover the still elusive function of NG2 cells and their synapses in the brain, rather than by its sensational but transient successes at upsetting the old order established by neuronal physiology. To participate in the effort to facilitate such a transition, here we propose a critical review of the latest findings in the field of NG2 cell physiology--discussing how they inform us on the possible function(s) of NG2 cells in the brain--and we present some personal views on new directions the field could benefit from in order to achieve lasting significance.
Figures
References
-
- Aguirre A, Dupree JL, Mangin JM, Gallo V. A functional role for EGFR signaling in myelination and remyelination. Nature Neurosci. 2007;10:990–1002. - PubMed
-
- Bakiri Y, Attwell D, Káradóttir R. Electrical signalling properties of oligodendrocyte precursor cells. Neuron Glia Biol. 2009;5:3–11. - PubMed
-
- Barres BA. The mystery and magic of glia: a perspective on their roles in health and disease. Neuron. 2008;60:430–440. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
