

Dot1 binding induces chromatin rearrangements by histone methylation-dependent and -independent mechanisms
- PMID: 21291527
- PMCID: PMC3038881
- DOI: 10.1186/1756-8935-4-2
Dot1 binding induces chromatin rearrangements by histone methylation-dependent and -independent mechanisms
Abstract
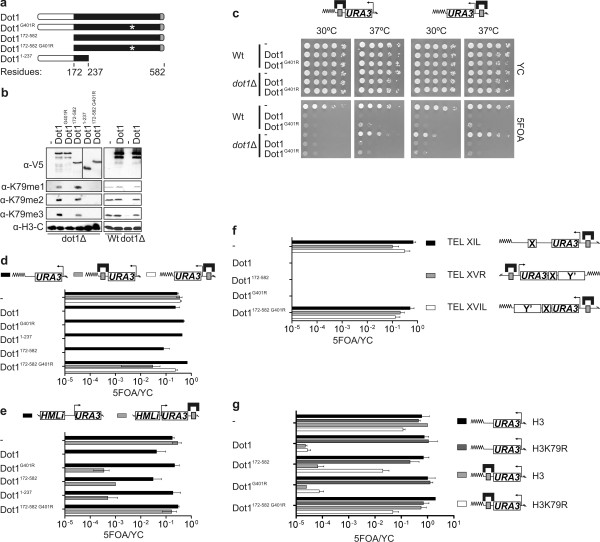
Background: Methylation of histone H3 lysine 79 (H3K79) by Dot1 is highly conserved among species and has been associated with both gene repression and activation. To eliminate indirect effects and examine the direct consequences of Dot1 binding and H3K79 methylation, we investigated the effects of targeting Dot1 to different positions in the yeast genome.
Results: Targeting Dot1 did not activate transcription at a euchromatic locus. However, chromatin-bound Dot1 derepressed heterochromatin-mediated gene silencing over a considerable distance. Unexpectedly, Dot1-mediated derepression was established by both a H3K79 methylation-dependent and a methylation-independent mechanism; the latter required the histone acetyltransferase Gcn5. By monitoring the localization of a fluorescently tagged telomere in living cells, we found that the targeting of Dot1, but not its methylation activity, led to the release of a telomere from the repressive environment at the nuclear periphery. This probably contributes to the activity-independent derepression effect of Dot1.
Conclusions: Targeting of Dot1 promoted gene expression by antagonizing gene repression through both histone methylation and chromatin relocalization. Our findings show that binding of Dot1 to chromatin can positively affect local gene expression by chromatin rearrangements over a considerable distance.
Figures



References
-
- Schulze JM, Jackson J, Nakanishi S, Gardner JM, Hentrich T, Haug J, Johnston M, Jaspersen SL, Kobor MS, Shilatifard A. Linking cell cycle to histone modifications: SBF and H2B monoubiquitination machinery and cell-cycle regulation of H3K79 dimethylation. Mol Cell. 2009;35:626–641. doi: 10.1016/j.molcel.2009.07.017. - DOI - PMC - PubMed
-
- Schubeler D, MacAlpine DM, Scalzo D, Wirbelauer C, Kooperberg C, van Leeuwen F, Gottschling DE, O'Neill LP, Turner BM, Delrow J. et al. The histone modification pattern of active genes revealed through genome-wide chromatin analysis of a higher eukaryote. Genes Dev. 2004;18:1263–1271. doi: 10.1101/gad.1198204. - DOI - PMC - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
