

Complement and viral pathogenesis
- PMID: 21292294
- PMCID: PMC3073741
- DOI: 10.1016/j.virol.2010.12.045
Complement and viral pathogenesis
Abstract
The complement system functions as an immune surveillance system that rapidly responds to infection. Activation of the complement system by specific recognition pathways triggers a protease cascade, generating cleavage products that function to eliminate pathogens, regulate inflammatory responses, and shape adaptive immune responses. However, when dysregulated, these powerful functions can become destructive and the complement system has been implicated as a pathogenic effector in numerous diseases, including infectious diseases. This review highlights recent discoveries that have identified critical roles for the complement system in the pathogenesis of viral infection.
Copyright © 2010 Elsevier Inc. All rights reserved.
Figures
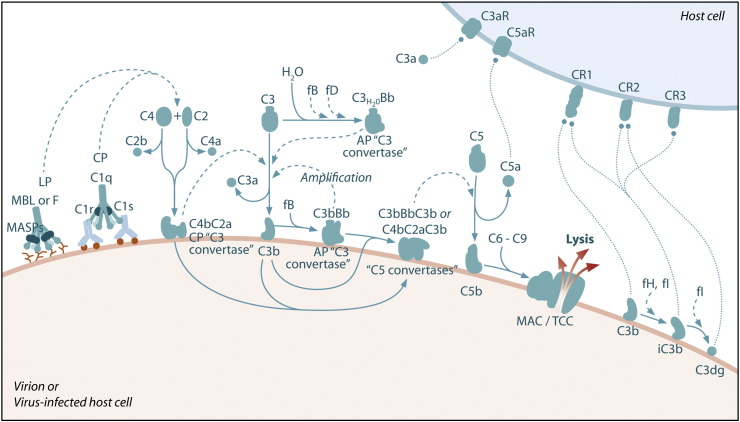
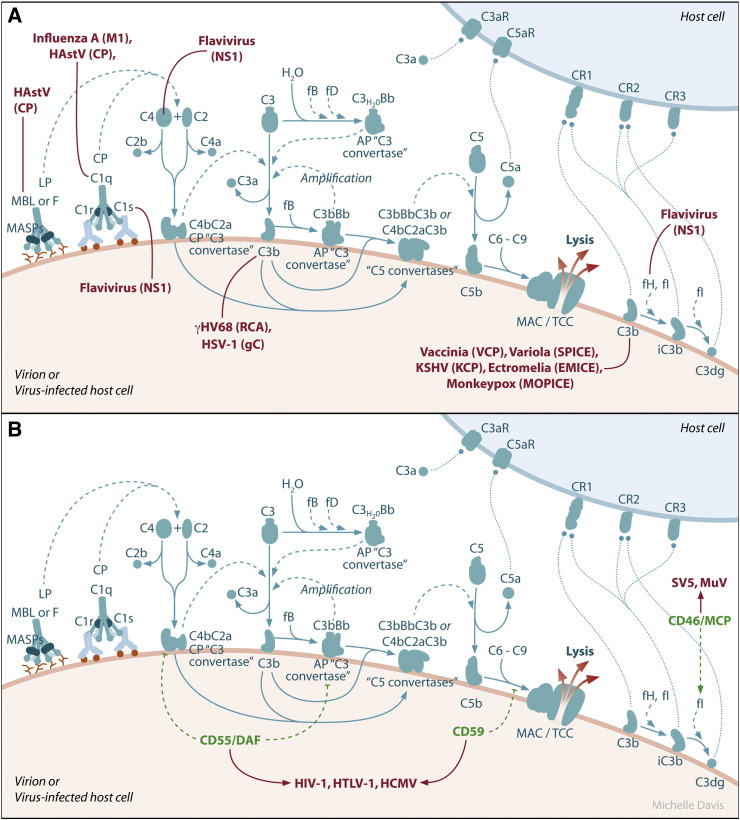
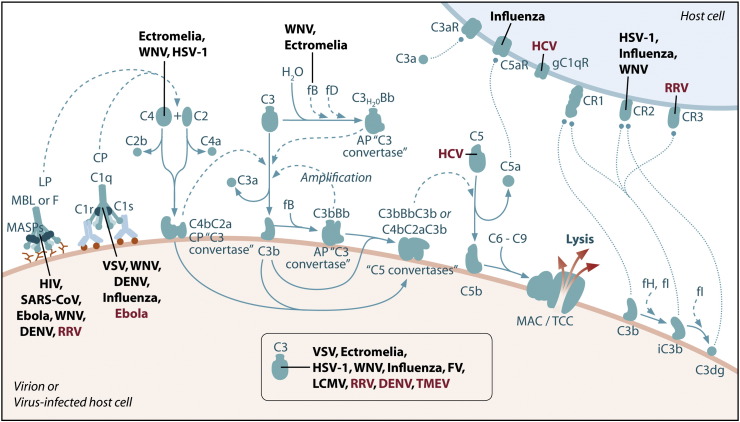
References
-
- Acioli-Santos B., Segat L., Dhalia R., Brito C.A., Braga-Neto U.M., Marques E.T., Crovella S. MBL2 gene polymorphisms protect against development of thrombocytopenia associated with severe dengue phenotype. Hum. Immunol. 2008;69(2):122–128. - PubMed
-
- Alcon-LePoder S., Sivard P., Drouet M.T., Talarmin A., Rice C., Flamand M. Secretion of flaviviral non-structural protein NS1: from diagnosis to pathogenesis. Novartis Found. Symp. 2006;277:233–247. discussion 247-53. - PubMed
-
- Alexandre K.B., Gray E.S., Lambson B.E., Moore P.L., Choge I.A., Mlisana K., Karim S.S., McMahon J., O'Keefe B., Chikwamba R., Morris L. Mannose-rich glycosylation patterns on HIV-1 subtype C gp120 and sensitivity to the lectins, Griffithsin, Cyanovirin-N and Scytovirin. Virology. 2010;402(1):187–196. - PMC - PubMed
-
- Auffermann-Gretzinger S., Keeffe E.B., Levy S. Impaired dendritic cell maturation in patients with chronic, but not resolved, hepatitis C virus infection. Blood. 2001;97(10):3171–3176. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
