

Genetic predisposition directs breast cancer phenotype by dictating progenitor cell fate
- PMID: 21295272
- PMCID: PMC3050563
- DOI: 10.1016/j.stem.2010.12.007
Genetic predisposition directs breast cancer phenotype by dictating progenitor cell fate
Abstract
Women with inherited mutations in the BRCA1 gene have increased risk of developing breast cancer but also exhibit a predisposition for the development of aggressive basal-like breast tumors. We report here that breast epithelial cells derived from patients harboring deleterious mutations in BRCA1 (BRCA1(mut /+) give rise to tumors with increased basal differentiation relative to cells from BRCA1+/+ patients. Molecular analysis of disease-free breast tissues from BRCA1(mut /+) patients revealed defects in progenitor cell lineage commitment even before cancer incidence. Moreover, we discovered that the transcriptional repressor Slug is an important functional suppressor of human breast progenitor cell lineage commitment and differentiation and that it is aberrantly expressed in BRCA1(mut /+) tissues. Slug expression is necessary for increased basal-like phenotypes prior to and after neoplastic transformation. These findings demonstrate that the genetic background of patient populations, in addition to affecting incidence rates, significantly impacts progenitor cell fate commitment and, therefore, tumor phenotype.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
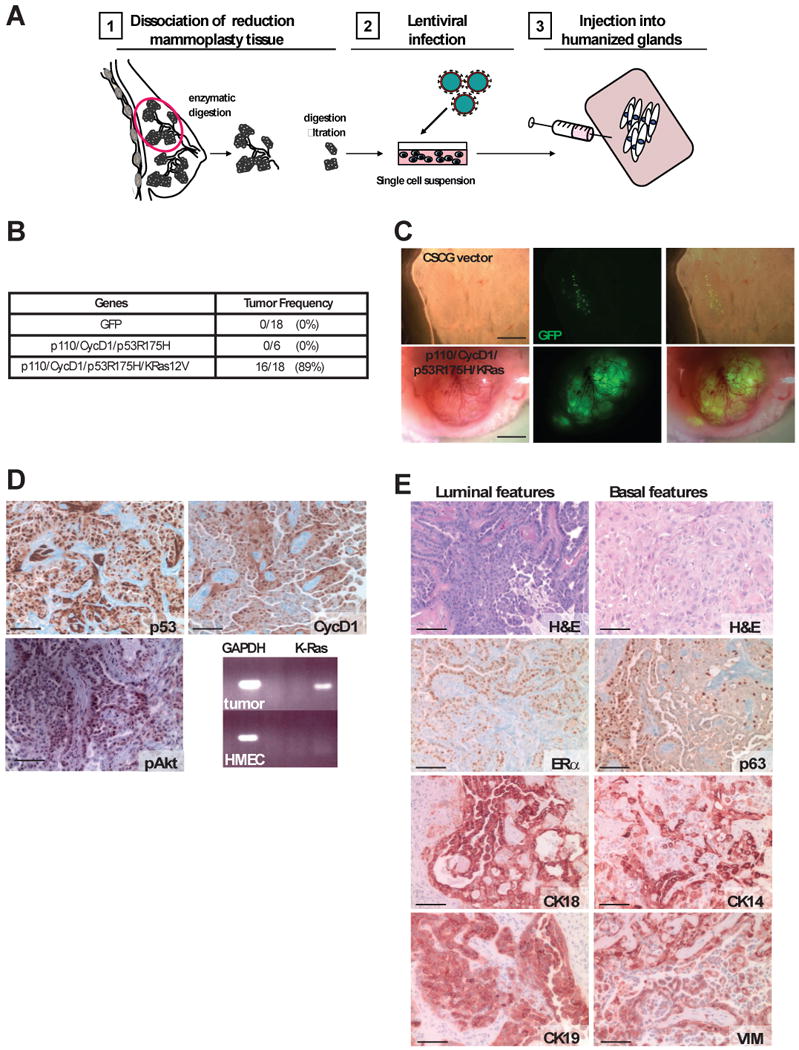
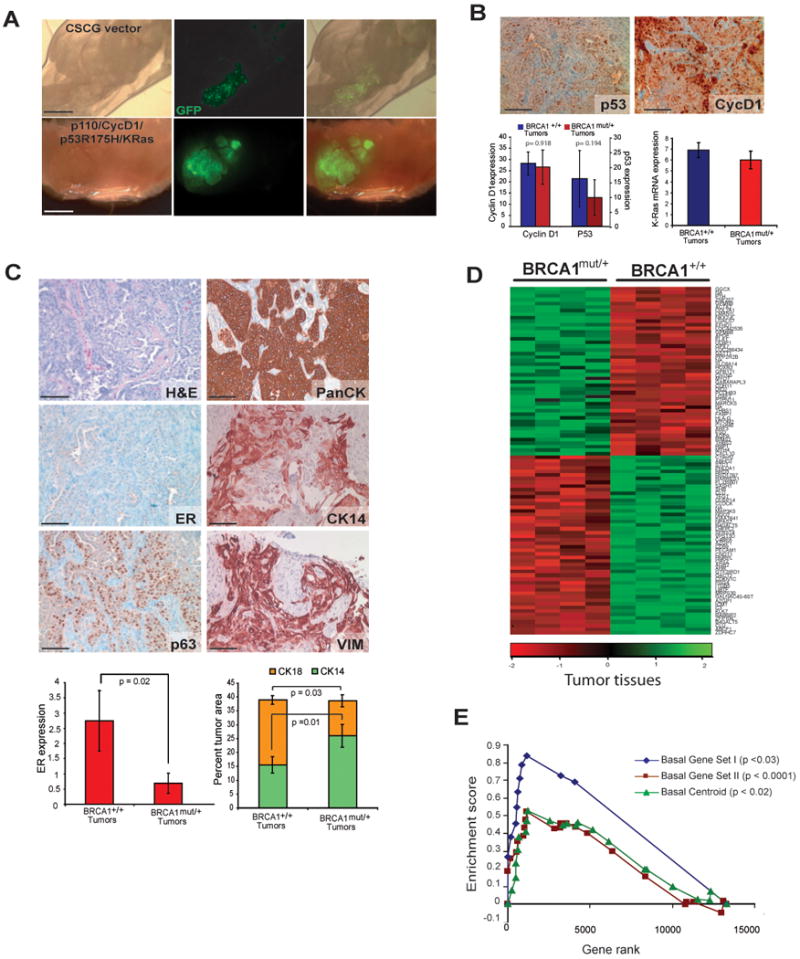
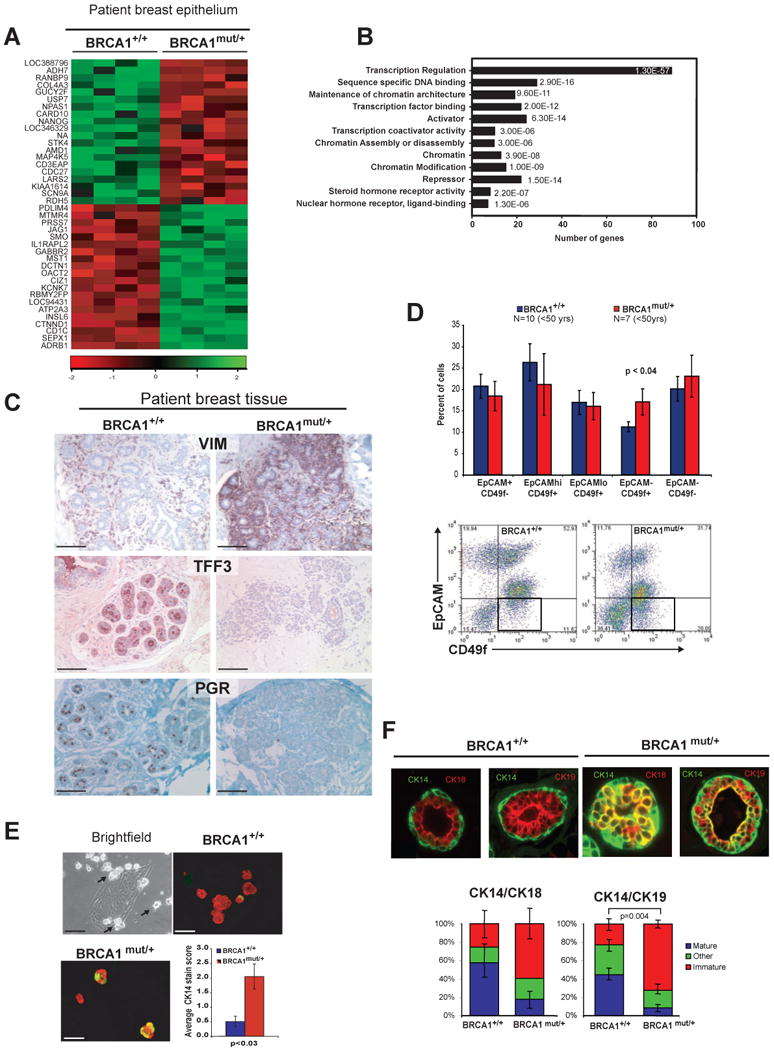
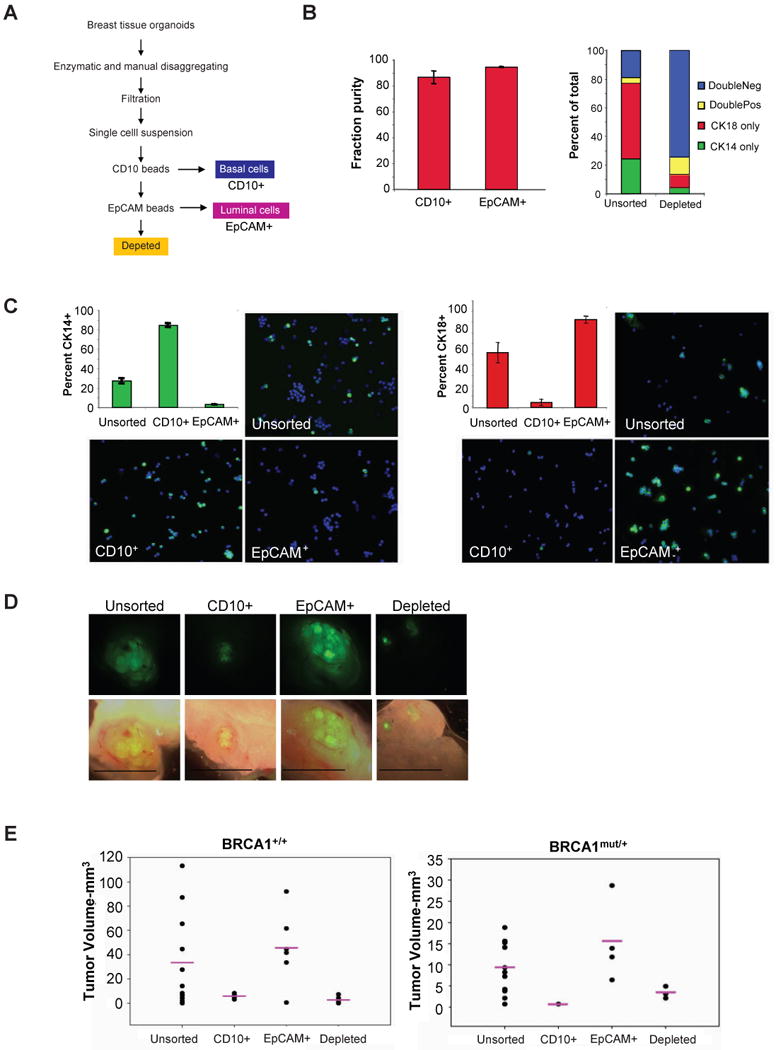
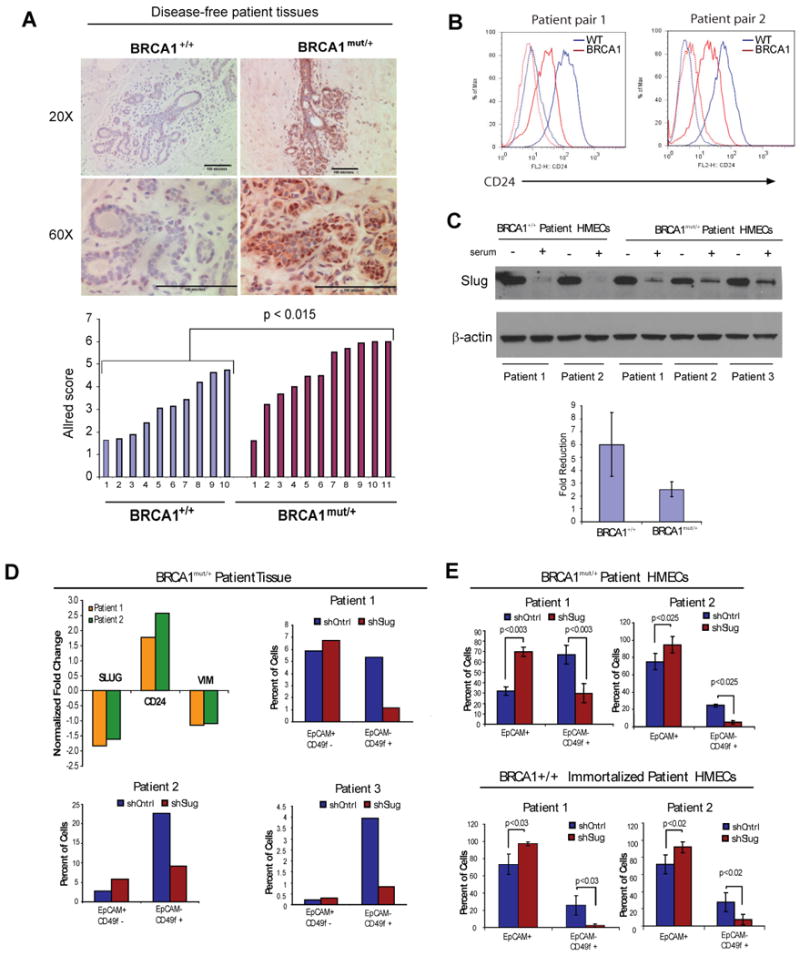
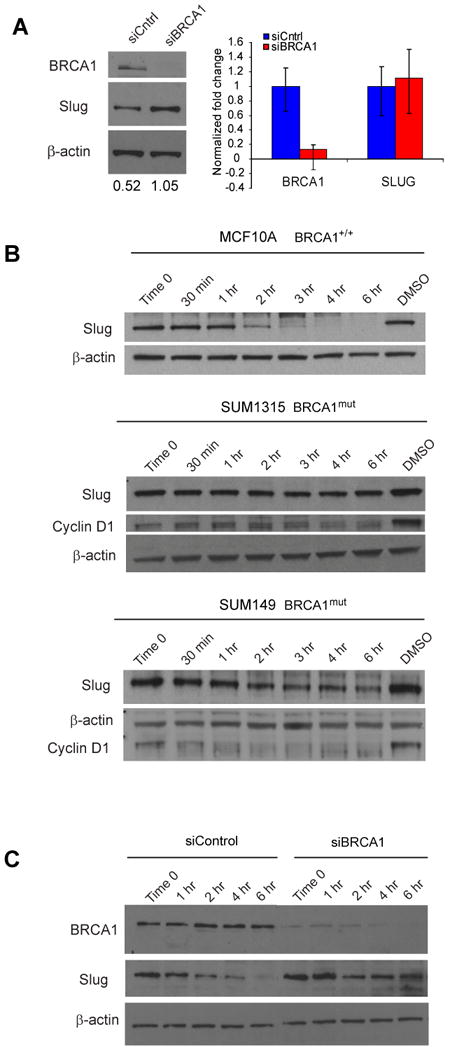
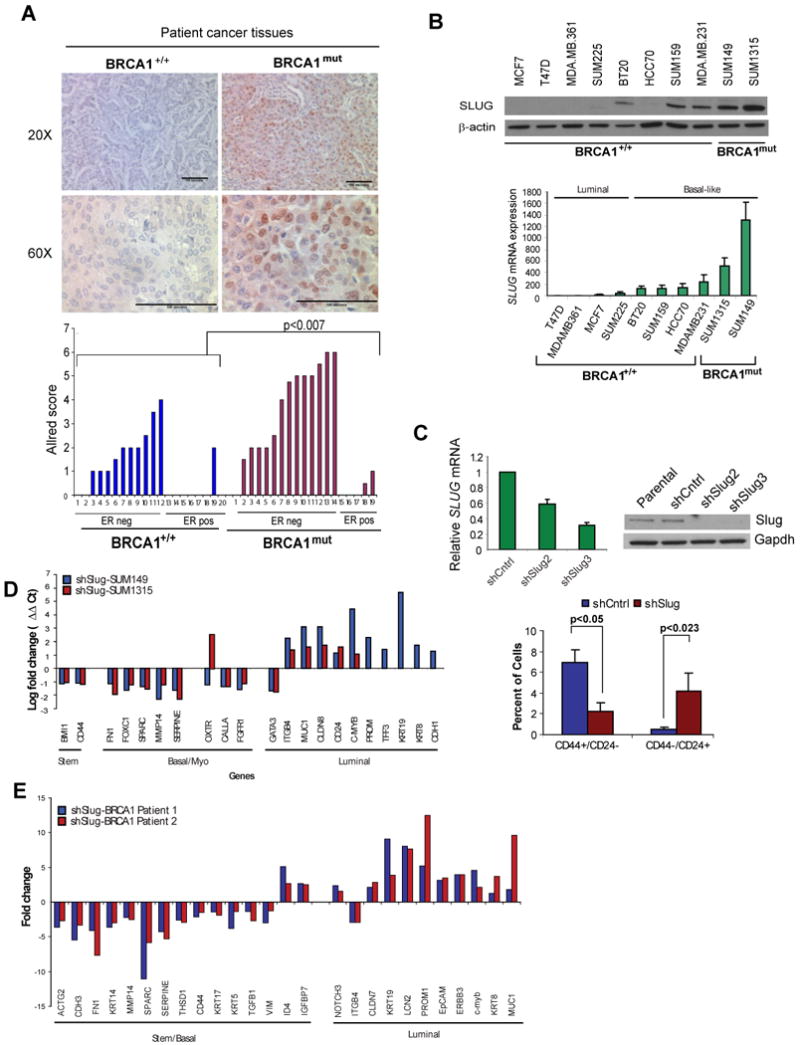
Comment in
-
A twist of cell fate.Cell Stem Cell. 2011 Feb 4;8(2):126-7. doi: 10.1016/j.stem.2011.01.004. Cell Stem Cell. 2011. PMID: 21295267
-
Breast cancer: Luminal cells with an identity crisis.Nat Rev Cancer. 2011 Mar;11(3):158. doi: 10.1038/nrc3030. Nat Rev Cancer. 2011. PMID: 21451553 No abstract available.
References
-
- Allinen M, Beroukhim R, Cai L, Brennan C, Lahti-Domenici J, Huang H, Porter D, Hu M, Chin L, Richardson A, Schnitt S, Sellers WR, Polyak K. Molecular characterization of the tumor microenvironment in breast cancer. Cancer Cell. 2004;6:17–32. - PubMed
-
- Arnes JB, Brunet JS, Stefansson I, Begin LR, Wong N, Chappuis PO, Akslen LA, Foulkes WD. Placental cadherin and the basal epithelial phenotype of BRCA1-related breast cancer. Clin Cancer Res. 2005;11:4003–4011. - PubMed
-
- Campbell IG, Russell SE, Choong DY, Montgomery KG, Ciavarella ML, Hooi CS, Cristiano BE, Pearson RB, Phillips WA. Mutation of the PIK3CA gene in ovarian and breast cancer. Cancer Res. 2004;64:7678–7681. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
