

Lineage analysis of micromere 4d, a super-phylotypic cell for Lophotrochozoa, in the leech Helobdella and the sludgeworm Tubifex
- PMID: 21295566
- PMCID: PMC3086575
- DOI: 10.1016/j.ydbio.2011.01.031
Lineage analysis of micromere 4d, a super-phylotypic cell for Lophotrochozoa, in the leech Helobdella and the sludgeworm Tubifex
Abstract
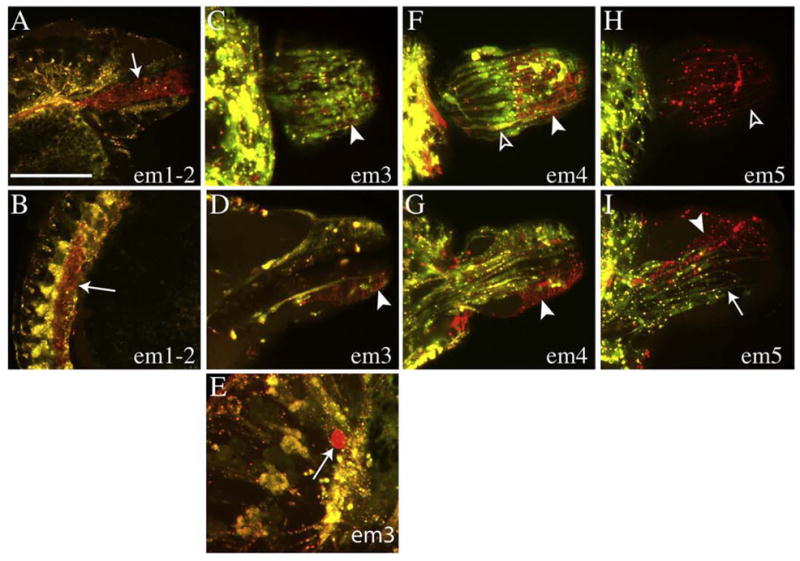
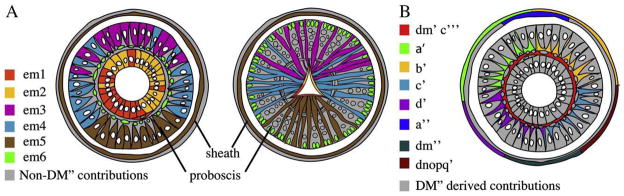
The super-phylum Lophotrochozoa contains the plurality of extant animal phyla and exhibits a corresponding diversity of adult body plans. Moreover, in contrast to Ecdysozoa and Deuterostomia, most lophotrochozoans exhibit a conserved pattern of stereotyped early divisions called spiral cleavage. In particular, bilateral mesoderm in most lophotrochozoan species arises from the progeny of micromere 4d, which is assumed to be homologous with a similar cell in the embryo of the ancestral lophotrochozoan, more than 650 million years ago. Thus, distinguishing the conserved and diversified features of cell fates in the 4d lineage among modern spiralians is required to understand how lophotrochozoan diversity has evolved by changes in developmental processes. Here we analyze cell fates for the early progeny of the bilateral daughters (M teloblasts) of micromere 4d in the leech Helobdella sp. Austin, a clitellate annelid. We show that the first six progeny of the M teloblasts (em1-em6) contribute five different sets of progeny to non-segmental mesoderm, mainly in the head and in the lining of the digestive tract. The latter feature, associated with cells em1 and em2 in Helobdella, is seen with the M teloblast lineage in a second clitellate species, the sludgeworm Tubifex tubifex and, on the basis of previously published work, in the initial progeny of the M teloblast homologs in molluscan species, suggesting that it may be an ancestral feature of lophotrochozoan development.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures










References
-
- Ackermann C, Dorresteijn A, Fischer A. Clonal domains in postlarval Platynereis dumerilii (Annelida: Polychaeta) J Morphol. 2005;266:258–280. - PubMed
-
- Anderson DT. Embryology and Phylogeny in Annelids and Arthropods. Pergamon; Oxford: 1973.
-
- Astrow S, Holton B, Weisblat DA. Centrifugation redistributes factors determining cleavage patterns in leech embryos. Dev Biol. 1987;120:270–283. - PubMed
-
- Bissen ST, Weisblat DA. The durations and compositions of cell cycles in embryos of the leech, Helobdella triserialis. Development. 1989;106:105–118. - PubMed
-
- Bouche M, Biagianti-Risbourge S, Vernet G. A light and scanning electron microscope study of the morphology of the chaetae of Tubifex tubifex in a non-polluted medium. Hydrobiologia. 1997;411:39–44.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
