

Non-proteolytic functions of calpain-3 in sarcoplasmic reticulum in skeletal muscles
- PMID: 21295580
- PMCID: PMC3056149
- DOI: 10.1016/j.jmb.2011.01.057
Non-proteolytic functions of calpain-3 in sarcoplasmic reticulum in skeletal muscles
Abstract
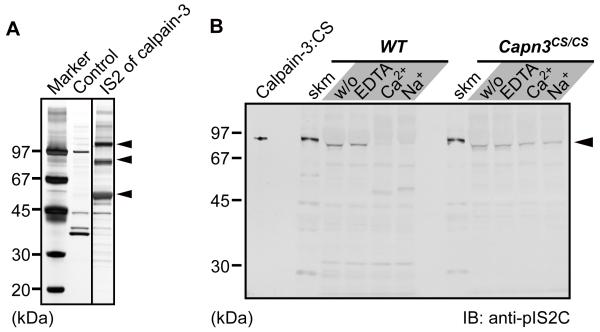
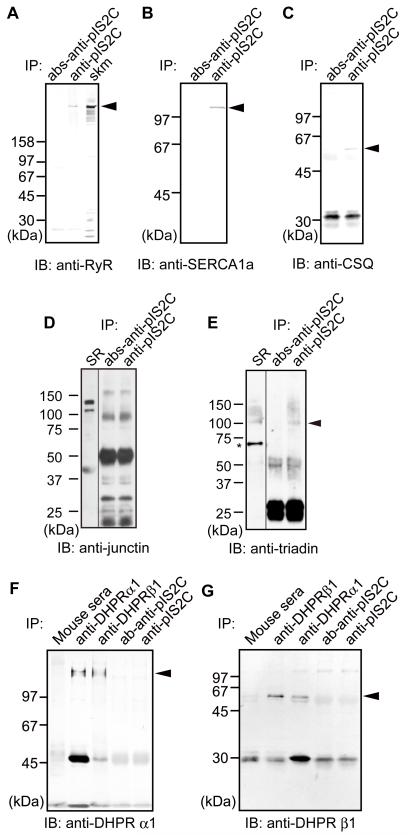
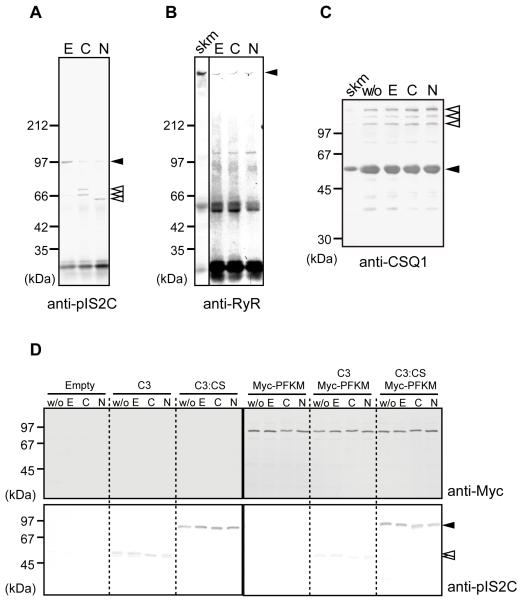
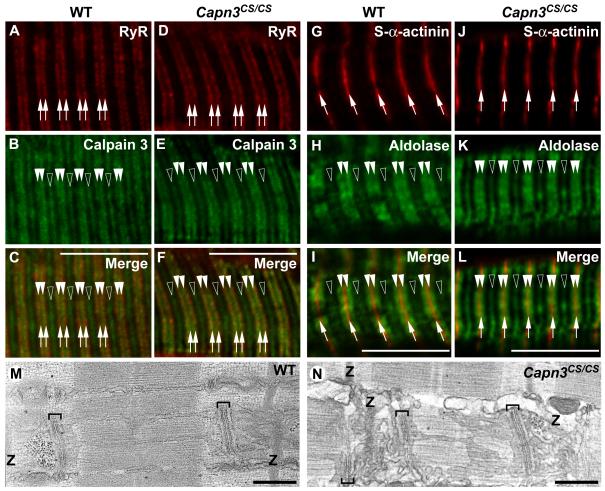
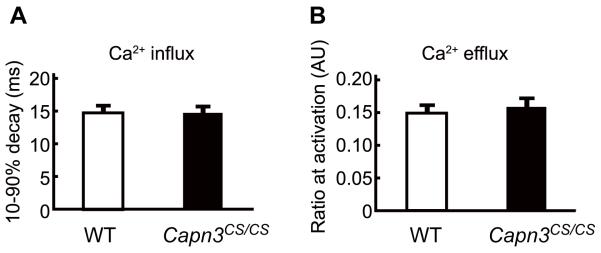
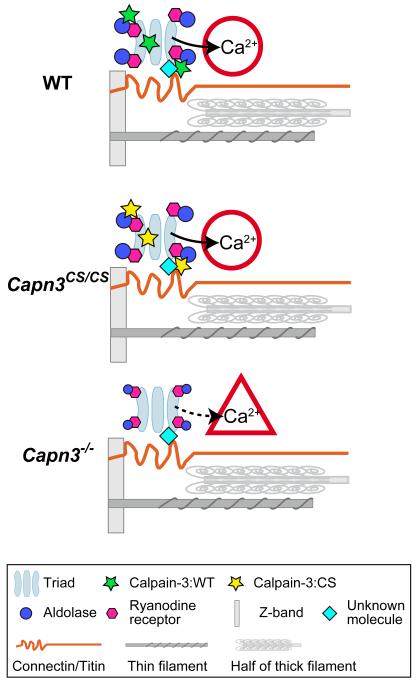
Mutations in CAPN3/Capn3, which codes for skeletal muscle-specific calpain-3/p94 protease, are responsible for limb-girdle muscular dystrophy type 2A. Using "knock-in" (referred to as Capn3(CS/CS)) mice, in which the endogenous calpain-3 is replaced with a mutant calpain-3:C129S, which is a proteolytically inactive but structurally intact calpain-3, we demonstrated in our previous studies that loss of calpain-3 protease activity causes muscular dystrophy [Ojima, K. et al. (2010) J. Clin. Invest. 120, 2672-2683]. However, compared to Capn3-null (Capn3(-/-)) mice, Capn3(CS/CS) mice showed less severe dystrophic symptoms. This suggests that calpain-3 also has a non-proteolytic function. This study aimed to elucidate the non-proteolytic functions of calpain-3 through comparison of Capn3(CS/CS) mice with Capn3(-/-) mice. We found that calpain-3 is a component of the sarcoplasmic reticulum (SR), and that calpain-3 interacts with, but does not proteolyze, typical SR components such as ryanodine receptor and calsequestrin. Furthermore, Capn3(CS/CS) mice showed that the nonenzymatic role of calpain-3 is required for proper Ca(2+) efflux from the SR to cytosol during muscle contraction. These results indicate that calpain-3 functions as a nonenzymatic element for the Ca(2+) efflux machinery in the SR, rather than as a protease. Thus, defects in the nonenzymatic function of calpain-3 must also be involved in the pathogenesis of limb-girdle muscular dystrophy type 2A.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Goll DE, Thompson VF, Li H, Wei W, Cong J. The calpain system. Physiol. Rev. 2003;83:731–801. - PubMed
-
- Suzuki K, Hata S, Kawabata Y, Sorimachi H. Structure, activation, and biology of calpain. Diabetes. 2004;53:S12–18. - PubMed
-
- Sorimachi H, Imajoh-Ohmi S, Emori Y, Kawasaki H, Ohno S, Minami Y, Suzuki K. Molecular cloning of a novel mammalian calcium-dependent protease distinct from both m- and mu-types. Specific expression of the mRNA in skeletal muscle. J. Biol. Chem. 1989;264:20106–20111. - PubMed
-
- Richard I, Broux O, Allamand V, Fougerousse F, Chiannilkulchai N, Bourg N, Brenguier L, Devaud C, Pasturaud P, Roudaut C, Hillaire D, Passos-Bueno M-R, Zats M, Tischfield JA, Fardeau M, Jackson CE, Cohen D, Beckmann JS. Mutations in the proteolytic enzyme calpain 3 cause limb-girdle muscular dystrophy type 2A. Cell. 1995;81:27–40. - PubMed
-
- Kramerova I, Kudryashova E, Tidball JG, Spencer MJ. Null mutation of calpain 3 (p94) in mice causes abnormal sarcomere formation in vivo and in vitro. Hum. Mol. Genet. 2004;13:1373–1388. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
