

Functional and mechanistic diversity of distal transcription enhancers
- PMID: 21295696
- PMCID: PMC3742076
- DOI: 10.1016/j.cell.2011.01.024
Functional and mechanistic diversity of distal transcription enhancers
Erratum in
- Cell. 2011 Mar 4;144(5):825
Abstract
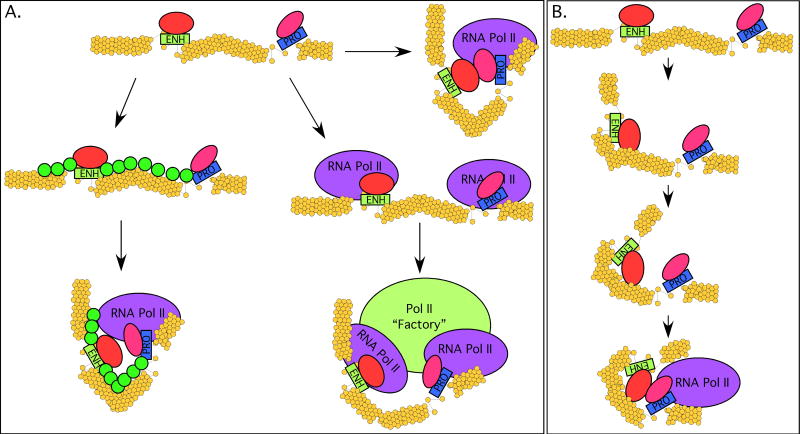
Biological differences among metazoans and between cell types in a given organism arise in large part due to differences in gene expression patterns. Gene-distal enhancers are key contributors to these expression patterns, exhibiting both sequence diversity and cell type specificity. Studies of long-range interactions indicate that enhancers are often important determinants of nuclear organization, contributing to a general model for enhancer function that involves direct enhancer-promoter contact. However, mechanisms for enhancer function are emerging that do not fit solely within such a model, suggesting that enhancers as a class of DNA regulatory element may be functionally and mechanistically diverse.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures


References
-
- Amano T, Sagai T, Tanabe H, Mizushina Y, Nakazawa H, Shiroishi T. Chromosomal dynamics at the Shh locus: limb bud-specific differential regulation of competence and active transcription. Dev Cell. 2009;16:47–57. - PubMed
-
- Andrulis ED, Neiman AM, Zappulla DC, Sternglanz R. Perinuclear localization of chromatin facilitates transcriptional silencing. Nature. 1998;394:592–595. - PubMed
-
- Banerji J, Rusconi S, Schaffner W. Expression of a beta-globin gene is enhanced by remote SV40 DNA sequences. Cell. 1981;27:299–308. - PubMed
-
- Banerji J, Olson L, Schaffner W. A lymphocyte-specific cellular enhancer is located downstream of the joining region in immunoglobulin heavy chain genes. Cell. 1983;33:729–740. - PubMed
-
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K. High-resolution profiling of histone methylations in the human genome. Cell. 2007;129:823–837. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
