

Nuclear pore complex-a coat specifically tailored for the nuclear envelope
- PMID: 21296566
- PMCID: PMC3109177
- DOI: 10.1016/j.ceb.2011.01.002
Nuclear pore complex-a coat specifically tailored for the nuclear envelope
Abstract
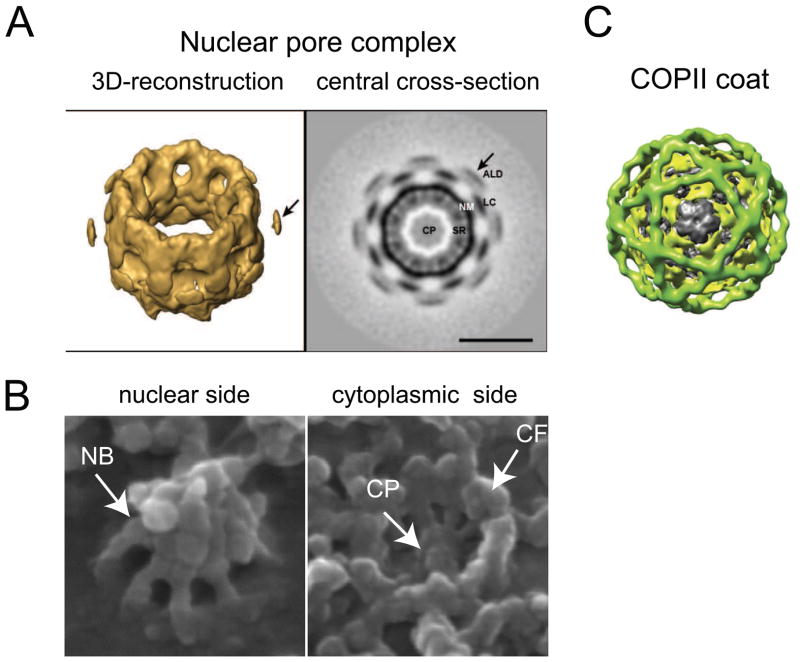
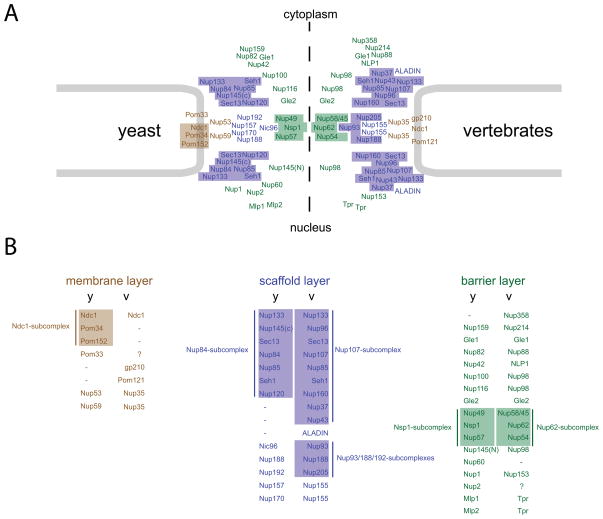
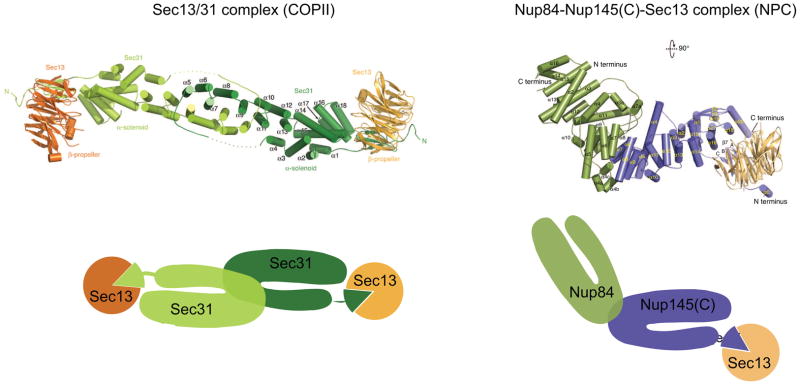
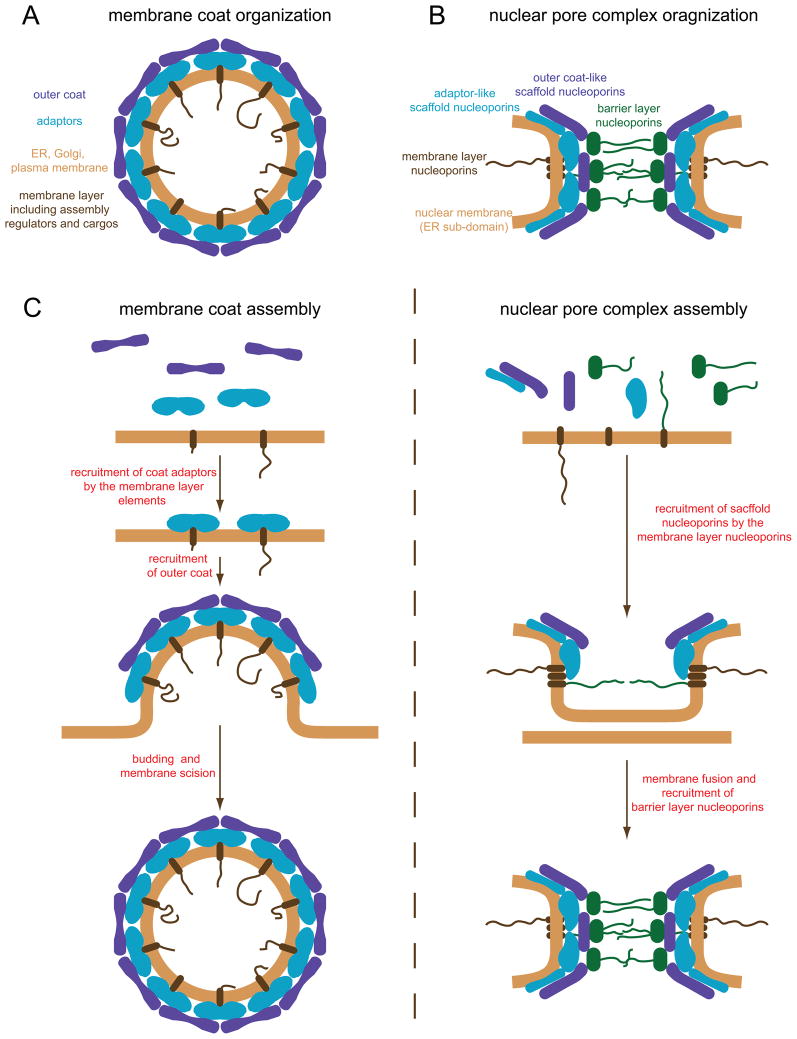
Nuclear pore complexes (NPCs) are highly selective transport gates that enable the bi-directional traffic of macromolecules across the nuclear envelope (NE). NPCs are located at the fusion pores between the inner and outer membranes of the NE and are built from a common set of ∼30 different proteins, nucleoporins. Remarkably, recent proteomic, bioinformatic, and structural studies have provided firm evidence that key structural nucleoporins share common ancestry with elements of coated vesicles, indicating an evolutionary link between these structures. This has provided novel insight into the origin of NPCs and may help us to better functionally characterize these fundamental components of eukaryotic cells.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Mans BJ, Anantharaman V, Aravind L, Koonin EV. Comparative genomics, evolution and origins of the nuclear envelope and nuclear pore complex. Cell Cycle. 2004;3:1612–1637. - PubMed
-
- DeGrasse JA, DuBois KN, Devos D, Siegel TN, Sali A, Field MC, Rout MP, Chait BT. Evidence for a shared nuclear pore complex architecture that is conserved from the last common eukaryotic ancestor. Mol Cell Proteomics. 2009;8:2119–2130. This paper describes the proteomic analysis of NPCs from Trypanosoma brucei, a highly divergent eukaryote, evolutionarily distant from species whose NPCs have been analyzed previously. The authors find that in spite of low sequence homology the majority of experimentally identified nucleoporins in trypanosomes have clear orthologs in yeast and vertebrates. Intriguingly, NPCs from T. brucei contain both scaffold and barrier layer nucleoporins, but transmbrane nucleoporins were not identified. Together these results suggest that a common NPC core structure was already present in the last eukaryotic common ancestor. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
