

Identification and characterization of genomic regions on chromosomes 4 and 8 that control the rate of photosynthesis in rice leaves
- PMID: 21296764
- PMCID: PMC3060679
- DOI: 10.1093/jxb/erq387
Identification and characterization of genomic regions on chromosomes 4 and 8 that control the rate of photosynthesis in rice leaves
Abstract
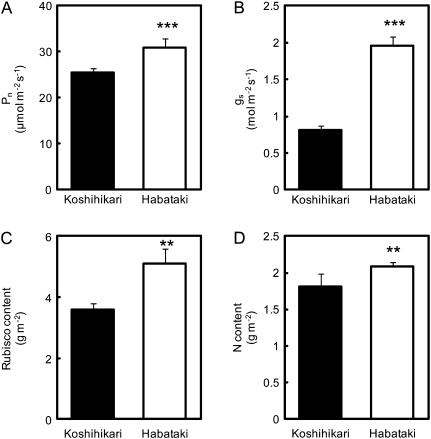
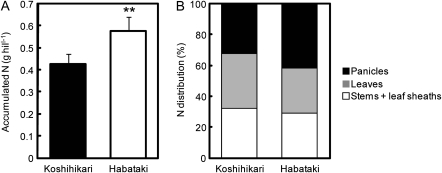
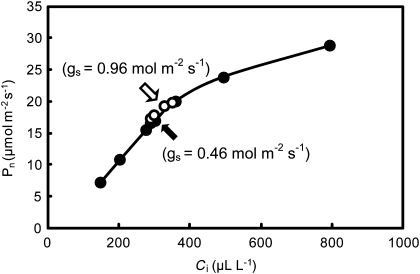
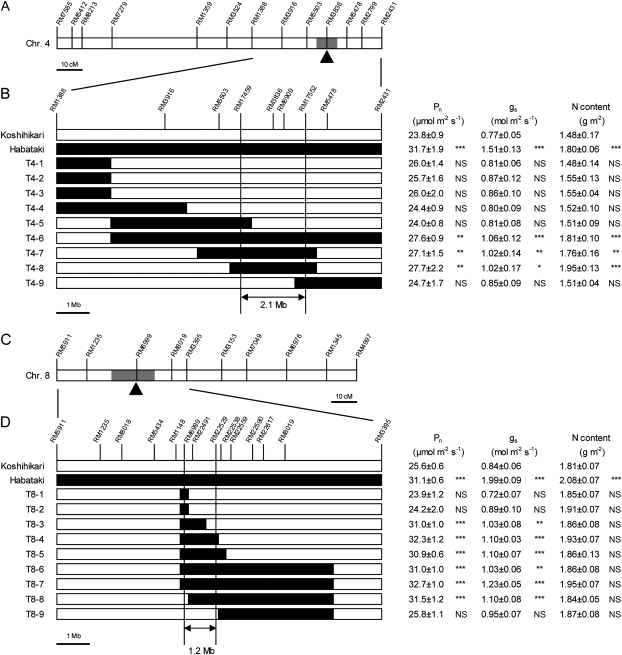
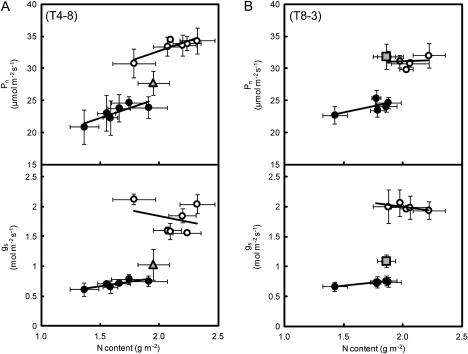
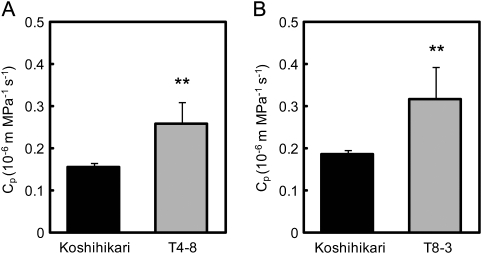
DNA marker-assisted selection appears to be a promising strategy for improving rates of leaf photosynthesis in rice. The rate of leaf photosynthesis was significantly higher in a high-yielding indica variety, Habataki, than in the most popular Japanese variety, Koshihikari, at the full heading stage as a result of the higher level of leaf nitrogen at the same rate of application of nitrogen and the higher stomatal conductance even when the respective levels of leaf nitrogen were the same. The higher leaf nitrogen content of Habataki was caused by the greater accumulation of nitrogen by plants. The higher stomatal conductance of Habataki was caused by the higher hydraulic conductance. Using progeny populations and selected lines derived from a cross between Koshihikari and Habataki, it was possible to identify the genomic regions responsible for the rate of photosynthesis within a 2.1 Mb region between RM17459 and RM17552 and within a 1.2 Mb region between RM6999 and RM22529 on the long arm of chromosome 4 and on the short arm of chromosome 8, respectively. The designated region on chromosome 4 of Habataki was responsible for both the increase in the nitrogen content of leaves and hydraulic conductance in the plant by increasing the root surface area. The designated region on chromosome 8 of Habataki was responsible for the increase in hydraulic conductance by increasing the root hydraulic conductivity. The results suggest that it may be possible to improve photosynthesis in rice leaves by marker-assisted selection that focuses on these regions of chromosomes 4 and 8.
Figures
References
-
- Asanuma S, Nito N, Ookawa T, Hirasawa T. Yield, dry matter production and ecophysiological characteristics of rice cultivar, Habataki compared with cv. Sasanishiki. Japanese Journal of Crop Science. 2008;77:474–480.
-
- Asanuma S, Ookawa T, Kondo M, Yano M, Ando T, Hirasawa T. QTL analysis of water transport capacity in rice: resistance to water transport in the lines of Habataki chromosome segment substitution in Sasanishiki. Proceedings of the 2nd International Conference on Rice for the Future. 2007:237.
-
- Basten CJ, Weir BS, Zeng ZB. A reference manual and tutorial for QTL mapping. Raleigh, NC: North Carolina State University; 2002. QTL cartographer. Version 1.16.
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous
