

The neural substrates of rapid-onset Dystonia-Parkinsonism
- PMID: 21297628
- PMCID: PMC3430603
- DOI: 10.1038/nn.2753
The neural substrates of rapid-onset Dystonia-Parkinsonism
Abstract
Although dystonias are a common group of movement disorders, the mechanisms by which brain dysfunction results in dystonia are not understood. Rapid-onset Dystonia-Parkinsonism (RDP) is a hereditary dystonia caused by mutations in the ATP1A3 gene. Affected individuals can be free of symptoms for years, but rapidly develop persistent dystonia and Parkinsonism-like symptoms after a stressful experience. Using a mouse model, we found that an adverse interaction between the cerebellum and basal ganglia can account for the symptoms of these individuals. The primary instigator of dystonia was the cerebellum, whose aberrant activity altered basal ganglia function, which in turn caused dystonia. This adverse interaction between the cerebellum and basal ganglia was mediated through a di-synaptic thalamic pathway that, when severed, alleviated dystonia. Our results provide a unifying hypothesis for the involvement of cerebellum and basal ganglia in the generation of dystonia and suggest therapeutic strategies for the treatment of RDP.
Figures
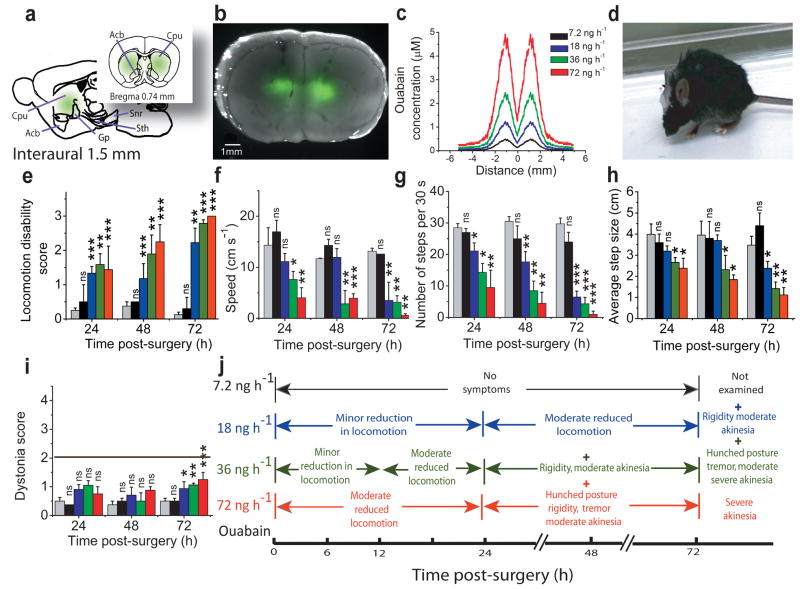

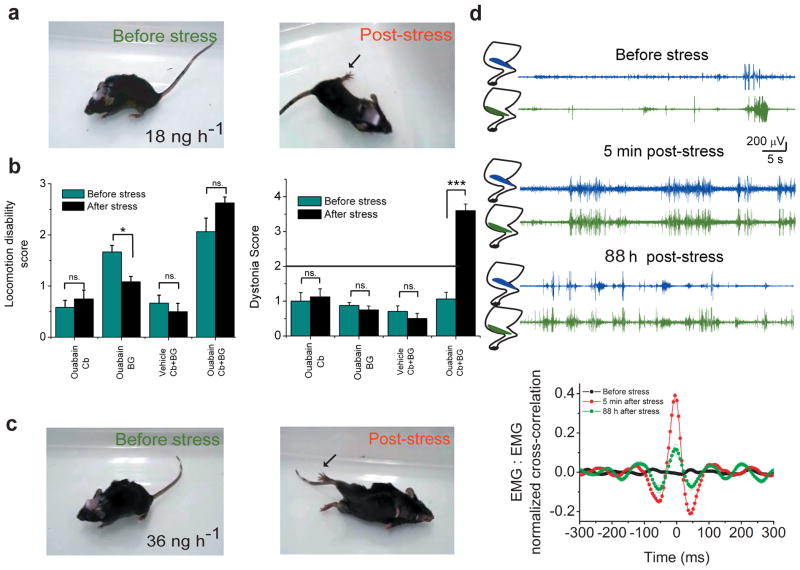
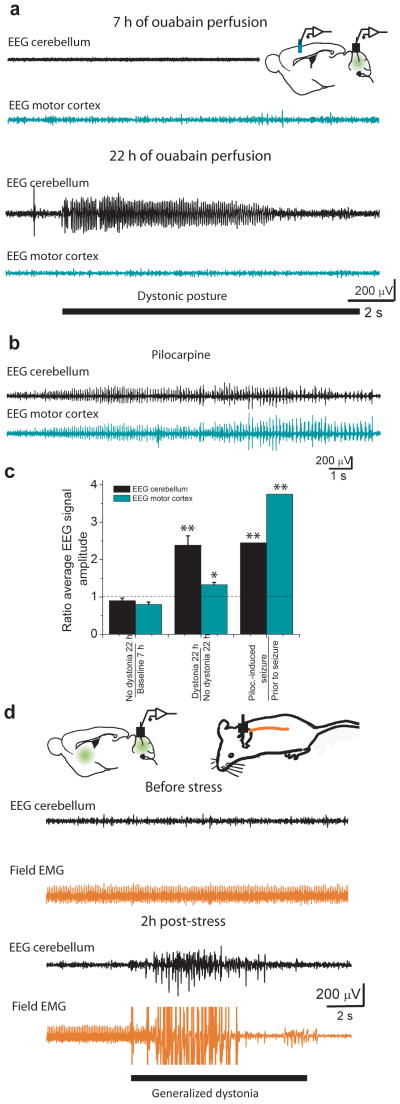
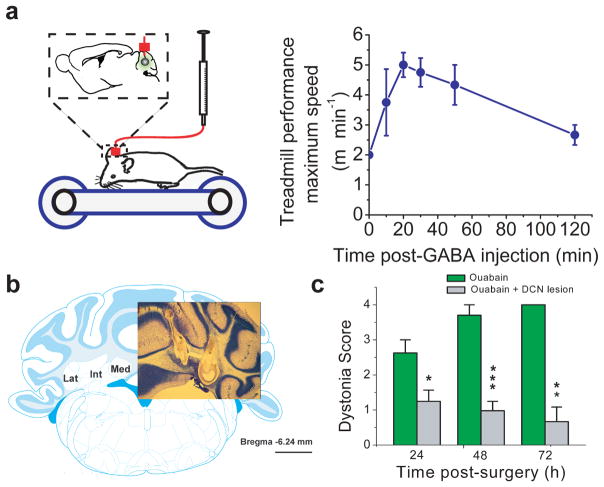

Comment in
-
Movement disorders: Cerebellum and basal ganglia interaction essential in RDP.Nat Rev Neurol. 2011 Apr;7(4):188. doi: 10.1038/nrneurol.2011.35. Epub 2011 Mar 15. Nat Rev Neurol. 2011. PMID: 21468120 No abstract available.
References
-
- Stacy MA. Handbook of Dystonia. Informa Health care; New York: 2006.
-
- Breakefield XO, et al. The pathophysiological basis of dystonias. Nat Rev Neurosci. 2008;9:222–234. - PubMed
-
- Muller U. The monogenic primary dystonias. Brain. 2009;132:2005–2025. - PubMed
-
- De Carvalho AP, et al. Mutations in the Na+/K+-ATPase alpha3 gene ATP1A3 are associated with rapid-onset dystonia parkinsonism. Neuron. 2004;43:169–175. - PubMed
-
- Brashear A, et al. The phenotypic spectrum of rapid-onset dystonia-parkinsonism (RDP) and mutations in the ATP1A3 gene. Brain. 2007;130:828–835. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
