

Differential in vitro effects of intravenous versus oral formulations of silibinin on the HCV life cycle and inflammation
- PMID: 21297992
- PMCID: PMC3030583
- DOI: 10.1371/journal.pone.0016464
Differential in vitro effects of intravenous versus oral formulations of silibinin on the HCV life cycle and inflammation
Abstract
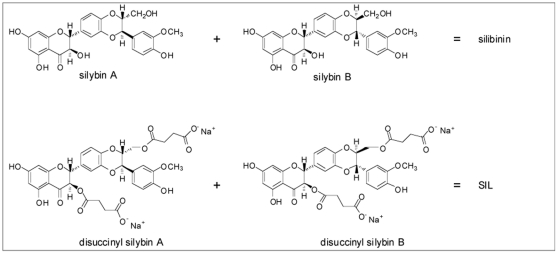
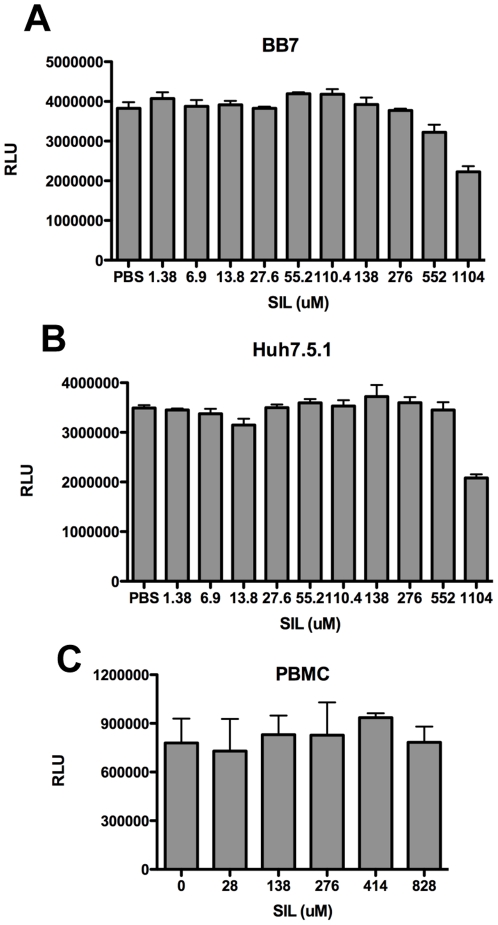
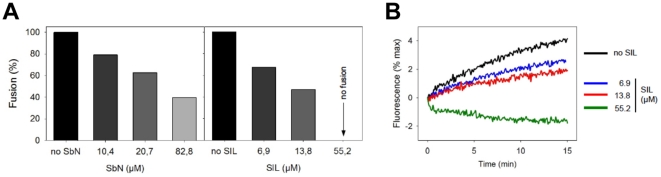
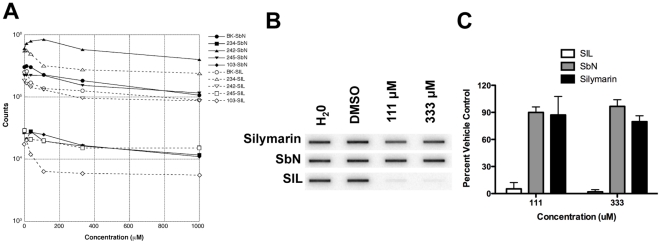
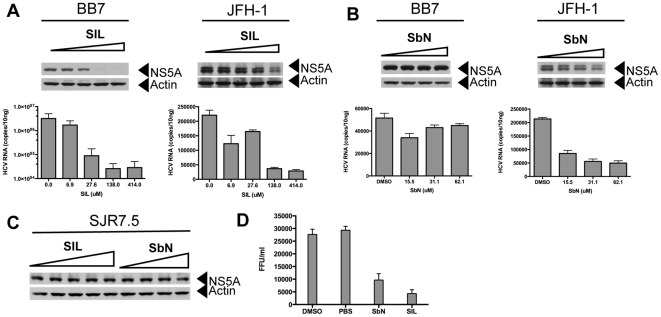
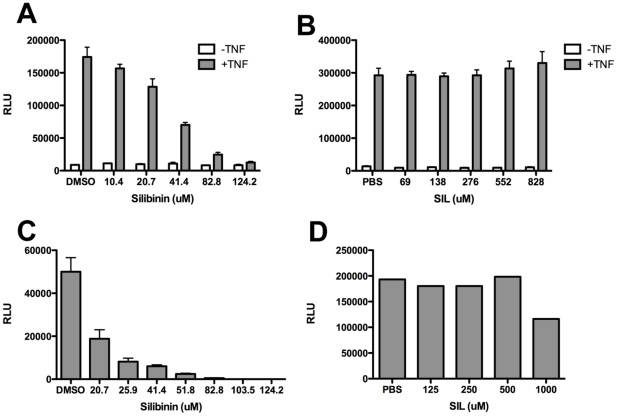
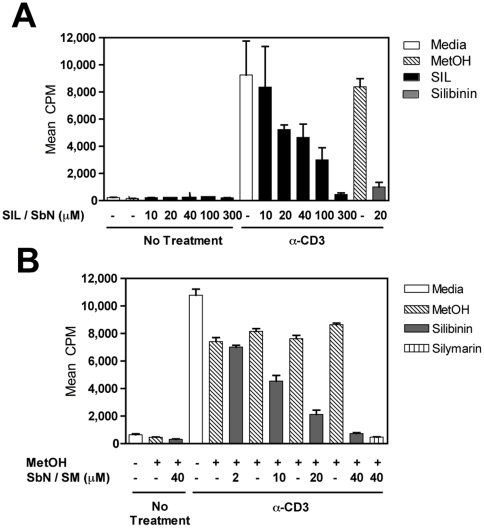
Silymarin prevents liver disease in many experimental rodent models, and is the most popular botanical medicine consumed by patients with hepatitis C. Silibinin is a major component of silymarin, consisting of the flavonolignans silybin A and silybin B, which are insoluble in aqueous solution. A chemically modified and soluble version of silibinin, SIL, has been shown to potently reduce hepatitis C virus (HCV) RNA levels in vivo when administered intravenously. Silymarin and silibinin inhibit HCV infection in cell culture by targeting multiple steps in the virus lifecycle. We tested the hepatoprotective profiles of SIL and silibinin in assays that measure antiviral and anti-inflammatory functions. Both mixtures inhibited fusion of HCV pseudoparticles (HCVpp) with fluorescent liposomes in a dose-dependent fashion. SIL inhibited 5 clinical genotype 1b isolates of NS5B RNA dependent RNA polymerase (RdRp) activity better than silibinin, with IC50 values of 40-85 µM. The enhanced activity of SIL may have been in part due to inhibition of NS5B binding to RNA templates. However, inhibition of the RdRps by both mixtures plateaued at 43-73%, suggesting that the products are poor overall inhibitors of RdRp. Silibinin did not inhibit HCV replication in subgenomic genotype 1b or 2a replicon cell lines, but it did inhibit JFH-1 infection. In contrast, SIL inhibited 1b but not 2a subgenomic replicons and also inhibited JFH-1 infection. Both mixtures inhibited production of progeny virus particles. Silibinin but not SIL inhibited NF-κB- and IFN-B-dependent transcription in Huh7 cells. However, both mixtures inhibited T cell proliferation to similar degrees. These data underscore the differences and similarities between the intravenous and oral formulations of silibinin, which could influence the clinical effects of this mixture on patients with chronic liver diseases.
Conflict of interest statement
Figures
References
-
- Perz JF, Armstrong GL, Farrington LA, Hutin YJ, Bell BP. The contributions of hepatitis B virus and hepatitis C virus infections to cirrhosis and primary liver cancer worldwide. J Hepatol. 2006;45:529–538. - PubMed
-
- Fried MW, Shiffman M, Sterling RK, Weinstein J, Crippin J, et al. A multicenter, randomized trial of daily high-dose interferon-alfa 2b for the treatment of chronic hepatitis c: pretreatment stratification by viral burden and genotype. Am J Gastroenterol. 2000;95:3225–3229. - PubMed
-
- Fried MW, Shiffman ML, Reddy KR, Smith C, Marinos G, et al. Peginterferon alfa-2a plus ribavirin for chronic hepatitis C virus infection. N Engl J Med. 2002;347:975–982. - PubMed
-
- Manns MP, McHutchison JG, Gordon SC, Rustgi VK, Shiffman M, et al. Peginterferon alfa-2b plus ribavirin compared with interferon alfa-2b plus ribavirin for initial treatment of chronic hepatitis C: a randomised trial. Lancet. 2001;358:958–965. - PubMed
-
- Kroll DJ, Shaw HS, Oberlies NH. Milk thistle nomenclature: why it matters in cancer research and pharmacokinetic studies. Integr Cancer Ther. 2007;6:110–119. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
