

Mammalian frataxin: an essential function for cellular viability through an interaction with a preformed ISCU/NFS1/ISD11 iron-sulfur assembly complex
- PMID: 21298097
- PMCID: PMC3027643
- DOI: 10.1371/journal.pone.0016199
Mammalian frataxin: an essential function for cellular viability through an interaction with a preformed ISCU/NFS1/ISD11 iron-sulfur assembly complex
Abstract
Background: Frataxin, the mitochondrial protein deficient in Friedreich ataxia, a rare autosomal recessive neurodegenerative disorder, is thought to be involved in multiple iron-dependent mitochondrial pathways. In particular, frataxin plays an important role in the formation of iron-sulfur (Fe-S) clusters biogenesis.
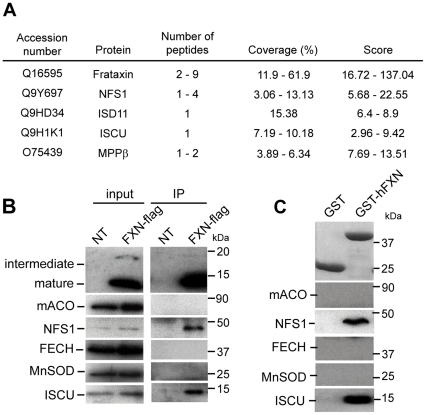
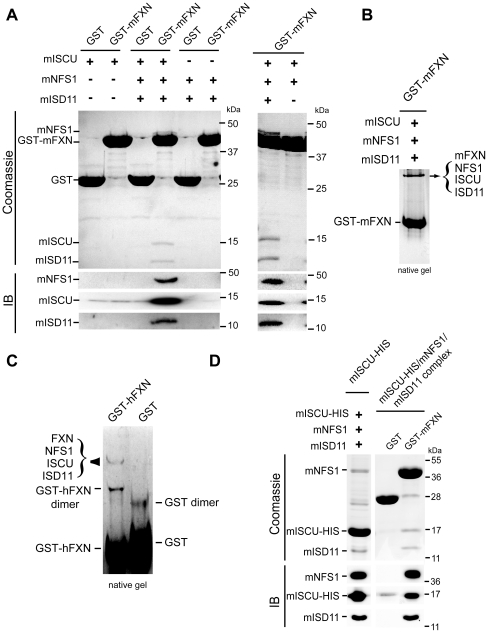
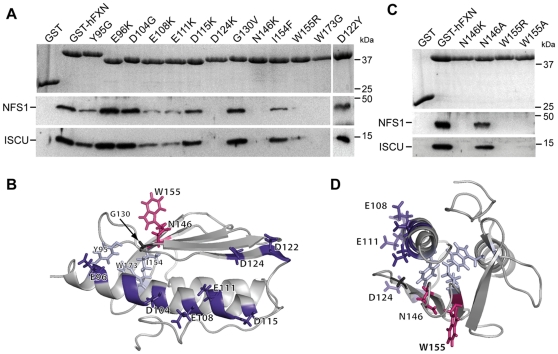
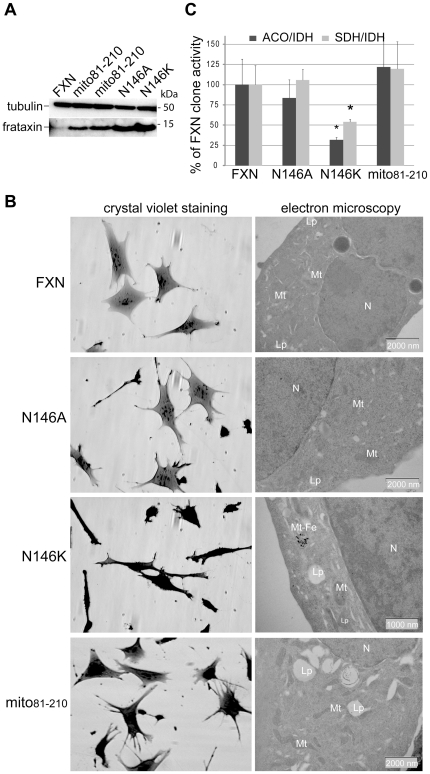
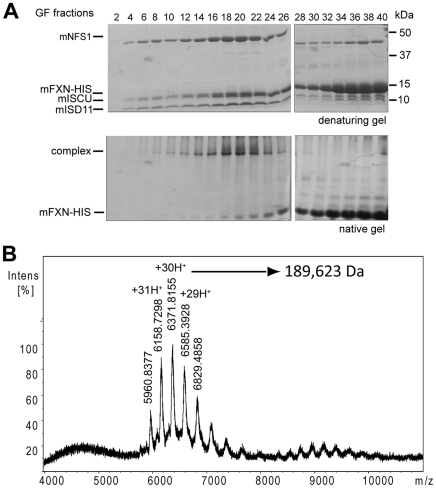
Methodology/principal findings: We present data providing new insights into the interactions of mammalian frataxin with the Fe-S assembly complex by combining in vitro and in vivo approaches. Through immunoprecipitation experiments, we show that the main endogenous interactors of a recombinant mature human frataxin are ISCU, NFS1 and ISD11, the components of the core Fe-S assembly complex. Furthermore, using a heterologous expression system, we demonstrate that mammalian frataxin interacts with the preformed core complex, rather than with the individual components. The quaternary complex can be isolated in a stable form and has a molecular mass of ≈190 kDa. Finally, we demonstrate that the mature human FXN(81-210) form of frataxin is the essential functional form in vivo.
Conclusions/significance: Our results suggest that the interaction of frataxin with the core ISCU/NFS1/ISD11 complex most likely defines the essential function of frataxin. Our results provide new elements important for further understanding the early steps of de novo Fe-S cluster biosynthesis.
Conflict of interest statement
Figures
References
-
- Campuzano V, Montermini L, Molto MD, Pianese L, Cossee M, et al. Friedreich's ataxia: autosomal recessive disease caused by an intronic GAA triplet repeat expansion. Science. 1996;271:1423–1427. - PubMed
-
- Schmucker S, Puccio H. Understanding the molecular mechanisms of Friedreich's ataxia to develop therapeutic approaches. Hum Mol Genet. 2010;19:R103–110. - PubMed
-
- Pandolfo M, Pastore A. The pathogenesis of Friedreich ataxia and the structure and function of frataxin. J Neurol. 2009;256(Suppl 1):9–17. - PubMed
-
- Nair M, Adinolfi S, Pastore C, Kelly G, Temussi P, et al. Solution structure of the bacterial frataxin ortholog, CyaY: mapping the iron binding sites. Structure. 2004;12:2037–2048. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
