

Resident memory T cells (T(RM)) are abundant in human lung: diversity, function, and antigen specificity
- PMID: 21298112
- PMCID: PMC3027667
- DOI: 10.1371/journal.pone.0016245
Resident memory T cells (T(RM)) are abundant in human lung: diversity, function, and antigen specificity
Abstract
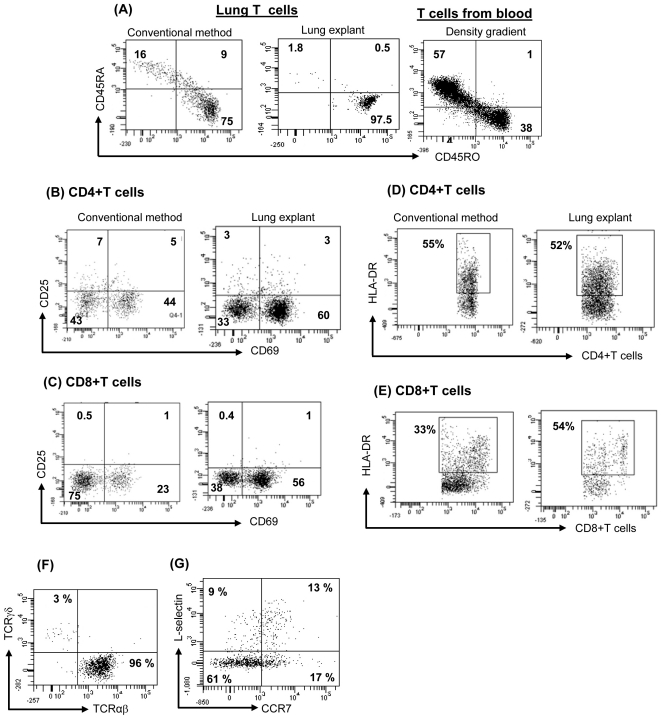
Recent studies have shown that tissue resident memory T cells (T(RM)) are critical to antiviral host defense in peripheral tissues. This new appreciation of T(RM) that reside in epithelial tissues and mediate host defense has been studied most extensively in skin: adult human skin contains large numbers of functional T(RM) that express skin specific markers. Indeed, more than twice as many T cells reside in skin as in peripheral blood. This T cell population has a diverse T cell receptor repertoire, and can produce a broad array of cytokines. More recently, we have begun to examine other epithelial tissues for the presence of resident T cells. In the present study, we asked whether analogous populations of resident T cells could be found in human lung. We were able to demonstrate abundant resident T cells in human lung-more than 10 billion T cells were present. Lung T cells were largely of the effector memory T cell (T(EM)) phenotype, though small numbers of central memory T cells (T(CM)) and T regulatory cells (T(reg)) could be identified. Lung T cells had a diverse T cell receptor repertoire and subsets produced IL-17, IL-4, IFNγ, as well as TNFα. A significant number of lung T(RM) CD4+Th cells produced more than one cytokine, identifying them as "multifunctional" Th1 type cells. Finally, lung T(RM), but not T(RM) resident to skin or T cells from blood, proliferated in response to influenza virus. This work suggests that normal human lung contains large numbers of T(RM) cells, and these cells are poised to respond to recall antigens previously encountered through lung mucosa. This population of T cells may contribute to the pathogenesis of asthma and other T cell mediated lung diseases.
Conflict of interest statement
Figures



References
-
- Luster AD, Alon R, von Andrian UH. Immune cell migration in inflammation: present and future therapeutic targets. Nat Immunol. 2005;6:1182–1190. - PubMed
-
- Robert C, Kupper TS. Inflammatory skin diseases, T cells, and immune surveillance. N Engl J Med. 1999;341:1817–1828. - PubMed
-
- Clark RA, Chong B, Mirchandani N, Brinster NK, Yamanaka K, et al. The vast majority of CLA+ T cells are resident in normal skin. J Immunol. 2006;176:4431–4439. - PubMed
-
- Clark RA, Chong BF, Mirchandani N, Yamanaka K, Murphy GF, et al. A novel method for the isolation of skin resident T cells from normal and diseased human skin. J Invest Dermatol. 2006;126:1059–1070. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
