

Potent and selective inhibitors of the inositol-requiring enzyme 1 endoribonuclease
- PMID: 21303903
- PMCID: PMC3069474
- DOI: 10.1074/jbc.M110.199737
Potent and selective inhibitors of the inositol-requiring enzyme 1 endoribonuclease
Abstract
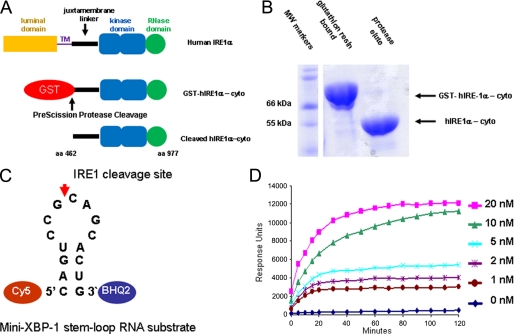
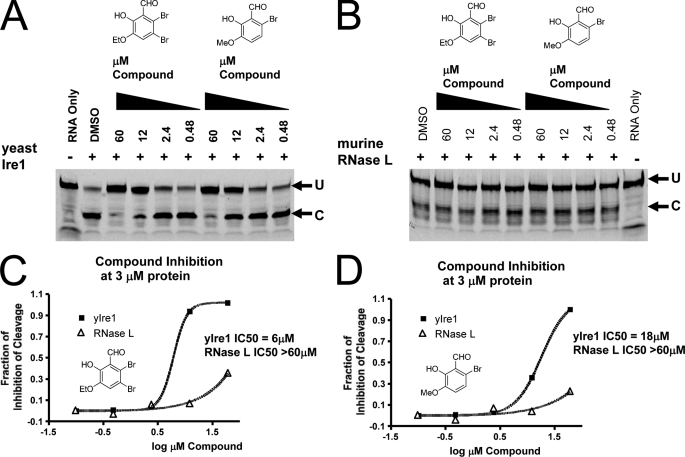
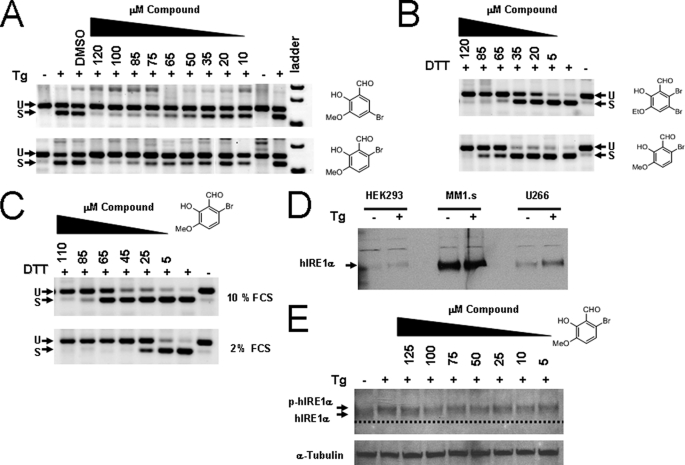
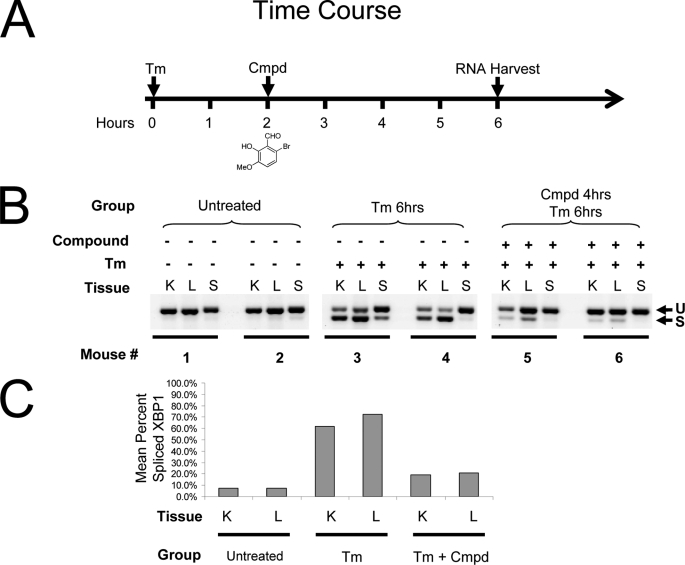
Inositol-requiring enzyme 1 (IRE1) is the most highly conserved signaling node of the unfolded protein response (UPR) and represents a potential therapeutic target for a number of diseases associated with endoplasmic reticulum stress. IRE1 activates the XBP-1 transcription factor by site-specific cleavage of two hairpin loops within its mRNA to facilitate its nonconventional splicing and alternative translation. We screened for inhibitors using a construct containing the unique cytosolic kinase and endoribonuclease domains of human IRE1α (hIRE1α-cyto) and a mini-XBP-1 stem-loop RNA as the substrate. One class compounds was salicylaldehyde analogs from the hydrolyzed product of salicylaldimines in the library. Salicylaldehyde analogs were active in inhibiting the site-specific cleavage of several mini-XBP-1 stem-loop RNAs in a dose-dependent manner. Salicyaldehyde analogs were also active in inhibiting yeast Ire1 but had little activity inhibiting RNase L or the unrelated RNases A and T1. Kinetic analysis revealed that one potent salicylaldehyde analog, 3-ethoxy-5,6-dibromosalicylaldehyde, is a non-competitive inhibitor with respect to the XBP-1 RNA substrate. Surface plasmon resonance studies confirmed this compound bound to IRE1 in a specific, reversible and dose-dependent manner. Salicylaldehydes inhibited XBP-1 splicing induced pharmacologically in human cells. These compounds also blocked transcriptional up-regulation of known XBP-1 targets as well as mRNAs targeted for degradation by IRE1. Finally, the salicylaldehyde analog 3-methoxy-6-bromosalicylaldehyde strongly inhibited XBP-1 splicing in an in vivo model of acute endoplasmic reticulum stress. To our knowledge, salicylaldehyde analogs are the first reported specific IRE1 endoribonuclease inhibitors.
Figures



Similar articles
-
Structure of the Ire1 autophosphorylation complex and implications for the unfolded protein response.EMBO J. 2011 Mar 2;30(5):894-905. doi: 10.1038/emboj.2011.18. Epub 2011 Feb 11. EMBO J. 2011. PMID: 21317875 Free PMC article.
-
IRE1alpha controls cyclin A1 expression and promotes cell proliferation through XBP-1.Cell Stress Chaperones. 2010 Sep;15(5):497-508. doi: 10.1007/s12192-009-0163-4. Epub 2009 Dec 15. Cell Stress Chaperones. 2010. PMID: 20013084 Free PMC article.
-
Selective inhibition of unfolded protein response induces apoptosis in pancreatic cancer cells.Oncotarget. 2014 Jul 15;5(13):4881-94. doi: 10.18632/oncotarget.2051. Oncotarget. 2014. PMID: 24952679 Free PMC article.
-
The molecular mechanism and functional diversity of UPR signaling sensor IRE1.Life Sci. 2021 Jan 15;265:118740. doi: 10.1016/j.lfs.2020.118740. Epub 2020 Nov 11. Life Sci. 2021. PMID: 33188833 Review.
-
Structural and molecular bases to IRE1 activity modulation.Biochem J. 2021 Aug 13;478(15):2953-2975. doi: 10.1042/BCJ20200919. Biochem J. 2021. PMID: 34375386 Review.
Cited by
-
Transcriptional regulation of secretory capacity by bZip transcription factors.Front Biol (Beijing). 2015 Feb 1;10(1):28-51. doi: 10.1007/s11515-014-1338-7. Front Biol (Beijing). 2015. PMID: 25821458 Free PMC article.
-
Endoplasmic Reticulum Stress in the Diabetic Kidney, the Good, the Bad and the Ugly.J Clin Med. 2015 Apr 20;4(4):715-40. doi: 10.3390/jcm4040715. J Clin Med. 2015. PMID: 26239352 Free PMC article. Review.
-
The role and therapeutic implication of endoplasmic reticulum stress in inflammatory cancer transformation.Am J Cancer Res. 2022 May 15;12(5):2277-2292. eCollection 2022. Am J Cancer Res. 2022. PMID: 35693091 Free PMC article. Review.
-
Dual RNase activity of IRE1 as a target for anticancer therapies.J Cell Commun Signal. 2023 Dec;17(4):1145-1161. doi: 10.1007/s12079-023-00784-5. Epub 2023 Sep 18. J Cell Commun Signal. 2023. PMID: 37721642 Free PMC article. Review.
-
Structural Tailoring of a Novel Fluorescent IRE-1 RNase Inhibitor to Precisely Control Its Activity.J Med Chem. 2019 Jun 13;62(11):5404-5413. doi: 10.1021/acs.jmedchem.9b00269. Epub 2019 May 22. J Med Chem. 2019. PMID: 31083990 Free PMC article.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
