

Understanding ribosome assembly: the structure of in vivo assembled immature 30S subunits revealed by cryo-electron microscopy
- PMID: 21303937
- PMCID: PMC3062180
- DOI: 10.1261/rna.2509811
Understanding ribosome assembly: the structure of in vivo assembled immature 30S subunits revealed by cryo-electron microscopy
Abstract
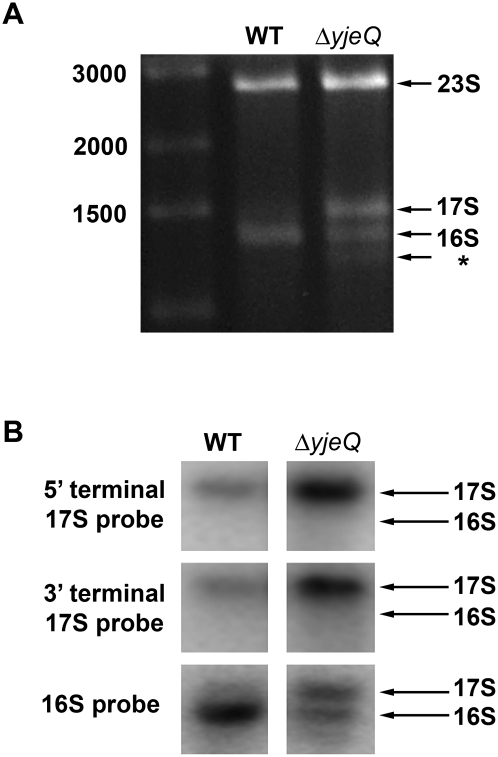
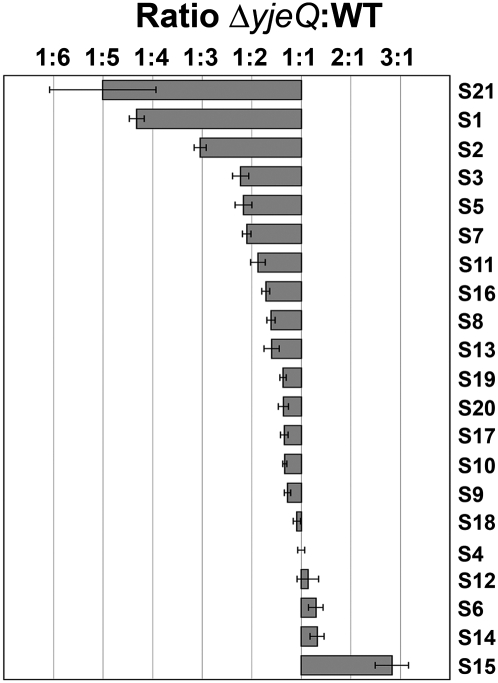
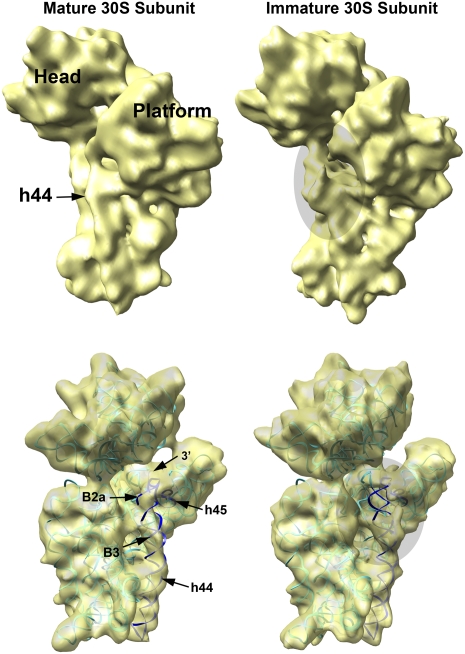
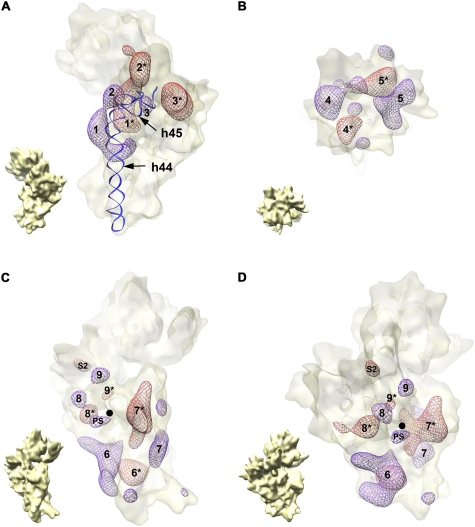
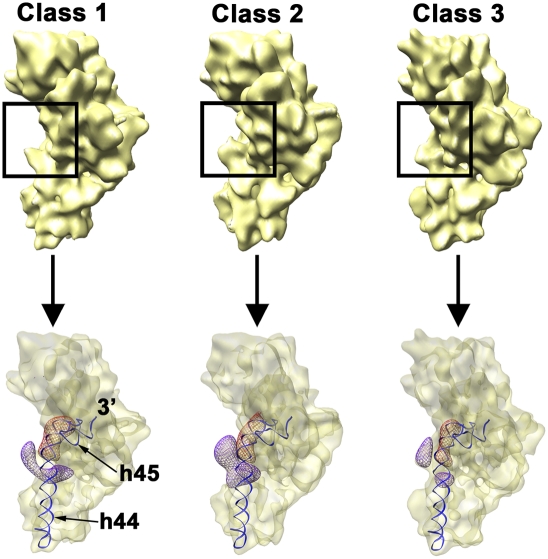
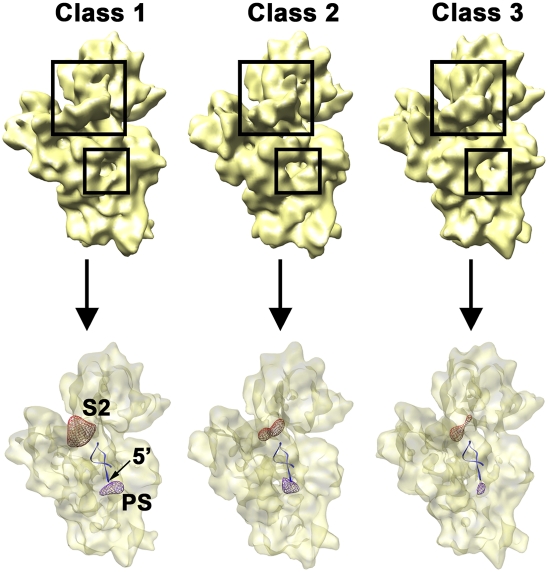
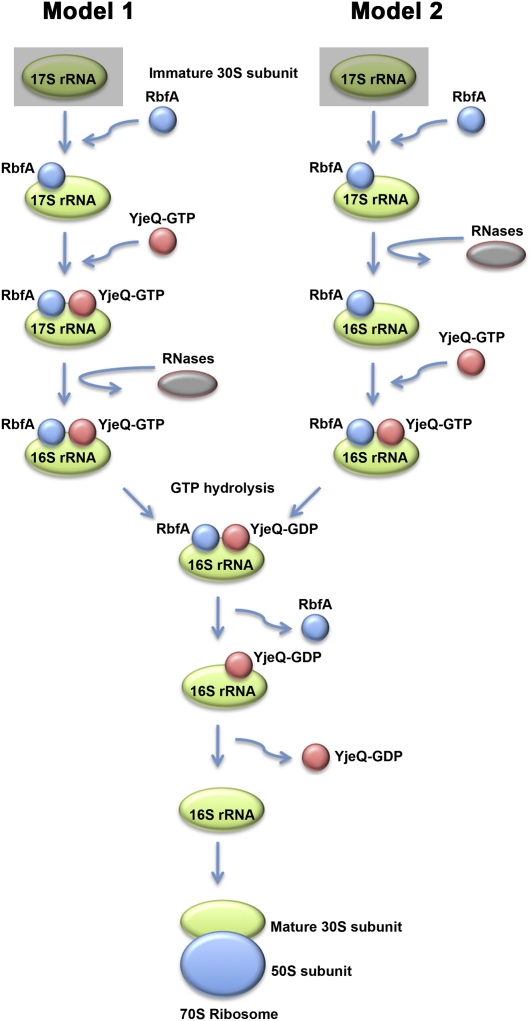
Four decades after early in vitro assembly studies demonstrated that ribosome assembly is a controlled process, our understanding of ribosome assembly is still incomplete. Just as structure determination has been so important to understanding ribosome function, so too will it be critical to sorting out the assembly process. Here, we used a viable deletion in the yjeQ gene, a recognized ribosome assembly factor, to isolate and structurally characterize immature 30S subunits assembled in vivo. These small ribosome subunits contained unprocessed 17S rRNA and lacked some late ribosomal proteins. Cryo-electron microscopy reconstructions revealed that the presence of precursor sequences in the rRNA induces a severe distortion in the 3' minor domain of the subunit involved in the decoding of mRNA and interaction with the large ribosome subunit. These findings suggest that rRNA processing events induce key local conformational changes directing the structure toward the mature assembly. We concluded that rRNA processing, folding, and the entry of tertiary r-proteins are interdependent events in the late stages of 30S subunit assembly. In addition, we demonstrate how studies of emerging assembly factors in ribosome biogenesis can help to elucidate the path of subunit assembly in vivo.
Figures
References
-
- Aebi U, Pollard TD 1987. A glow discharge unit to render electron microscope grids and other surfaces hydrophilic. J Electron Microsc Tech 7: 29–33 - PubMed
-
- Ban N, Nissen P, Hansen J, Moore PB, Steitz TA 2000. The complete atomic structure of the large ribosomal subunit at 2.4 Å resolution. Science 289: 905–920 - PubMed
-
- Borovinskaya MA, Pai RD, Zhang W, Schuwirth BS, Holton JM, Hirokawa G, Kaji H, Kaji A, Cate JH 2007. Structural basis for aminoglycoside inhibition of bacterial ribosome recycling. Nat Struct Mol Biol 14: 727–732 - PubMed
-
- Brown ED 2005. Conserved P-loop GTPases of unknown function in bacteria: an emerging and vital ensemble in bacterial physiology. Biochem Cell Biol 83: 738–746 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases