

2'-hydroxyflavanone inhibits proliferation, tumor vascularization and promotes normal differentiation in VHL-mutant renal cell carcinoma
- PMID: 21304051
- PMCID: PMC3066423
- DOI: 10.1093/carcin/bgr021
2'-hydroxyflavanone inhibits proliferation, tumor vascularization and promotes normal differentiation in VHL-mutant renal cell carcinoma
Abstract
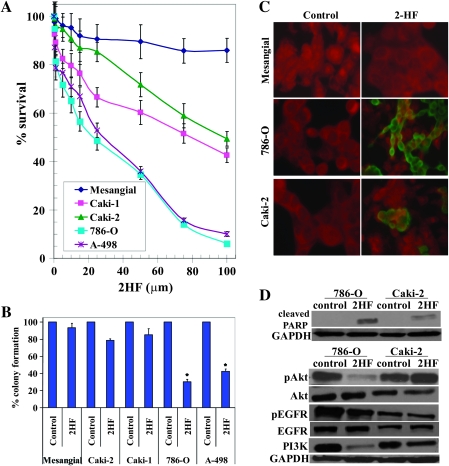
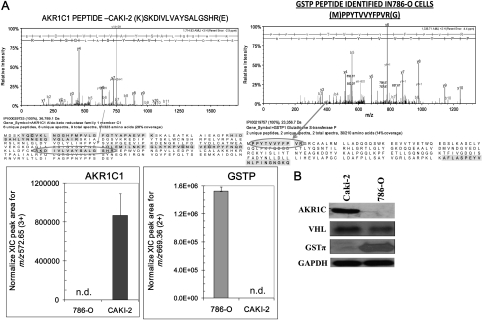
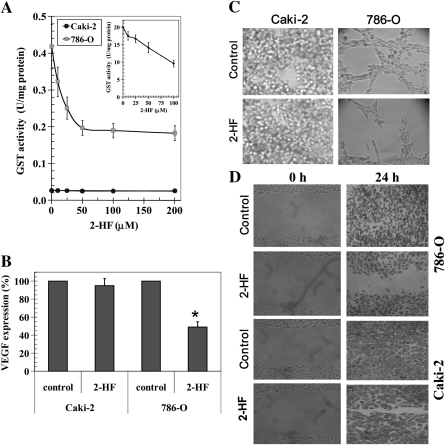
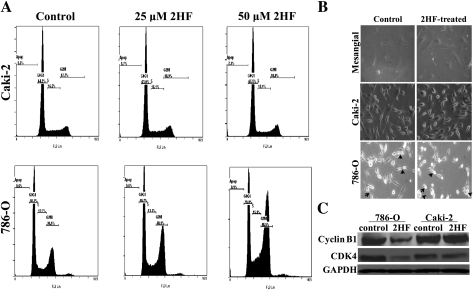
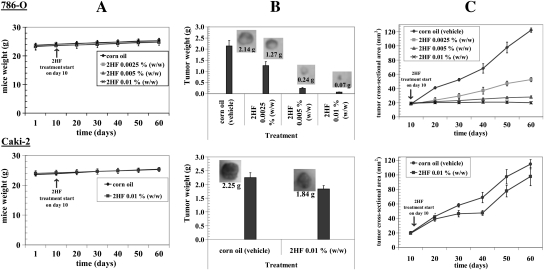
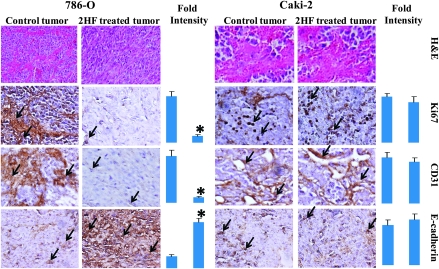
Renal cell carcinoma (RCC) is one of the top ten cancers prevalent in USA. Loss-of-function mutations in the von Hippel-Lindau (VHL) gene constitute an established risk factor contributing to 75% of total reported cases of RCC. Loss-of-VHL leads to a highly vascularized phenotype of renal tumors. Intake of citrus fruits has been proven to reduce the risk of RCC in multicenter international studies. Hence, we studied the effect of 2'-hydroxyflavanone (2HF), an active anticancer compound from oranges, in RCC. Our in vitro investigations revealed that 2HF suppresses VHL-mutant RCC to a significantly greater extent than VHL-wild-type RCC by inhibiting epidermal growth factor receptor signaling, which is increased due to VHL mutations in RCC. Our results also revealed for the first time, that 2HF inhibits glutathione S-transferase pi activity. 2HF reduced cyclin B1 and CDK4 levels and induced G2/M phase arrest in VHL-mutant RCC. Importantly, 2HF inhibited the angiogenesis in VHL-mutant RCC by decreasing vascular endothelial growth factor expression. Our in vivo studies in mice xenografts confirmed our in vitro results as evident by decreased levels of proliferation marker, Ki67 and angiogenic marker, CD31, in 2HF-treated mice xenografts of VHL-mutant RCC. 2HF also increased the expression of E-cadherin in VHL-mutant RCC, which would be of significance in restoring normal epithelial phenotype. Collectively, our in vitro and in vivo results revealed the potent antiproliferative, anti-angiogenic and prodifferentiation properties of 2HF in VHL-mutant RCC, sparing normal cells, which could have significant implications not only in the specific management of VHL-mutant RCC but also towards other VHL syndromes.
Figures
References
-
- Linehan WM, et al. Genetic basis of cancer of the kidney: disease-specific approaches to therapy. Clin. Cancer Res. 2004;10:6282S–6289S. - PubMed
-
- Atkins MB, et al. Innovations and challenges in renal cell carcinoma: summary statement from the Second Cambridge Conference. Clin. Cancer Res. 2007;13:667s–670s. - PubMed
-
- Kuroda N, et al. Review of renal oncocytoma with focus on clinical and pathobiological aspects. Histol. Histopathol. 2003;18:935–942. - PubMed
-
- Hunt JD, et al. Renal cell carcinoma in relation to cigarette smoking: meta-analysis of 24 studies. Int. J. Cancer. 2005;114:101–108. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
