

The Ufm1-activating enzyme Uba5 is indispensable for erythroid differentiation in mice
- PMID: 21304510
- PMCID: PMC3105337
- DOI: 10.1038/ncomms1182
The Ufm1-activating enzyme Uba5 is indispensable for erythroid differentiation in mice
Abstract
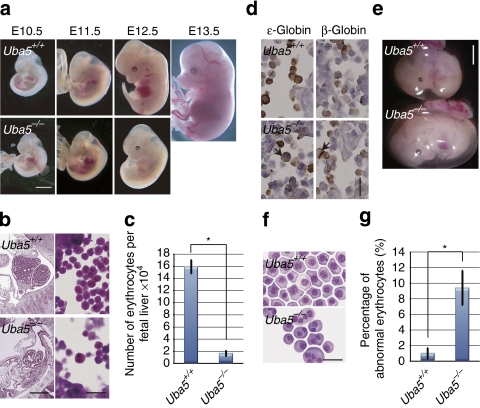
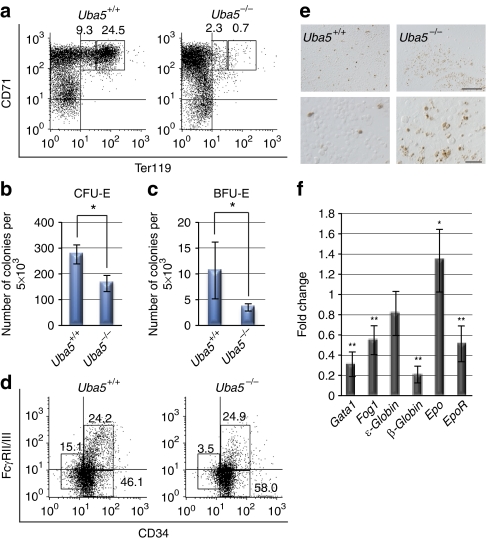
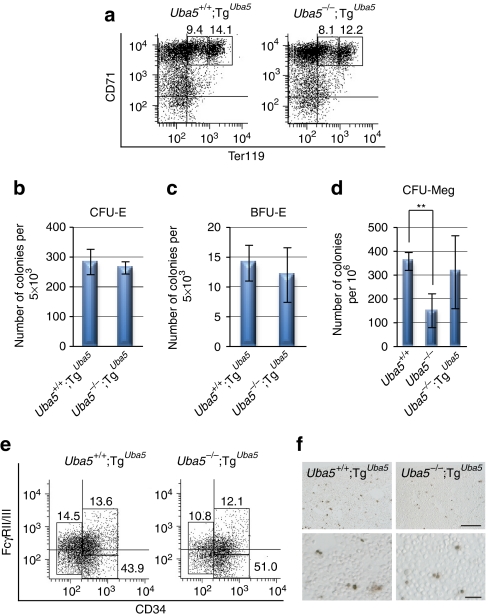
Post-translational protein modifications are systems designed to expand restricted genomic information through functional conversion of target molecules. Ubiquitin-like post-translational modifiers regulate numerous cellular events through their covalent linkages to target protein(s) by an enzymatic cascade analogous to ubiquitylation consisting of E1 (activating), E2 (conjugating) and E3 (ligating) enzymes. In this study, we report the essential role of Uba5, a specific activating enzyme for the ubiquitin-like modifier, Ufm1, in erythroid development. Mice lacking Uba5 exhibited severe anaemia, followed by death in utero. Although Uba5 was dispensable for the production of erythropoietin, its genetic loss led to impaired development of megakaryocyte and erythroid progenitors from common myeloid progenitors. Intriguingly, transgenic expression of Uba5 in the erythroid lineage rescued the Uba5-deficient embryos from anaemia and prolonged their survival, demonstrating the importance of Uba5 in cell-autonomous erythroid differentiation. Our results suggest that one of the ubiquitin-like protein modification systems, the Ufm1 system, is involved in the regulation of haematopoiesis.
Figures

References
-
- Hershko A. & Ciechanover A. The ubiquitin system. Annu. Rev. Biochem. 67, 425–479 (1998). - PubMed
-
- Goldberg A. L. Protein degradation and protection against misfolded or damaged proteins. Nature 426, 895–899 (2003). - PubMed
-
- Iwasaki H. & Akashi K. Myeloid lineage commitment from the hematopoietic stem cell. Immunity 26, 726–740 (2007). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
