

Early target cells of measles virus after aerosol infection of non-human primates
- PMID: 21304593
- PMCID: PMC3029373
- DOI: 10.1371/journal.ppat.1001263
Early target cells of measles virus after aerosol infection of non-human primates
Abstract
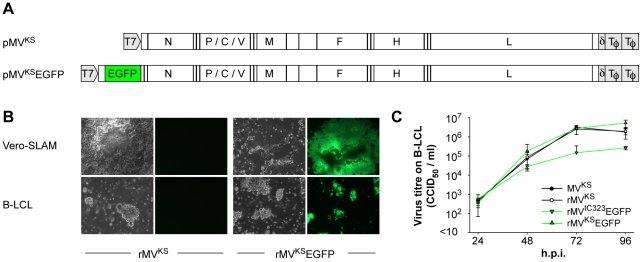
Measles virus (MV) is highly infectious, and has long been thought to enter the host by infecting epithelial cells of the respiratory tract. However, epithelial cells do not express signaling lymphocyte activation molecule (CD150), which is the high-affinity cellular receptor for wild-type MV strains. We have generated a new recombinant MV strain expressing enhanced green fluorescent protein (EGFP), based on a wild-type genotype B3 virus isolate from Khartoum, Sudan (KS). Cynomolgus macaques were infected with a high dose of rMV(KS)EGFP by aerosol inhalation to ensure that the virus could reach the full range of potential target cells throughout the entire respiratory tract. Animals were euthanized 2, 3, 4 or 5 days post-infection (d.p.i., n = 3 per time point) and infected (EGFP(+)) cells were identified at all four time points, albeit at low levels 2 and 3 d.p.i. At these earliest time points, MV-infected cells were exclusively detected in the lungs by fluorescence microscopy, histopathology and/or virus isolation from broncho-alveolar lavage cells. On 2 d.p.i., EGFP(+) cells were phenotypically typed as large mononuclear cells present in the alveolar lumen or lining the alveolar epithelium. One to two days later, larger clusters of MV-infected cells were detected in bronchus-associated lymphoid tissue (BALT) and in the tracheo-bronchial lymph nodes. From 4 d.p.i. onward, MV-infected cells were detected in peripheral blood and various lymphoid tissues. In spite of the possibility for the aerosolized virus to infect cells and lymphoid tissues of the upper respiratory tract, MV-infected cells were not detected in either the tonsils or the adenoids until after onset of viremia. These data strongly suggest that in our model MV entered the host at the alveolar level by infecting macrophages or dendritic cells, which traffic the virus to BALT or regional lymph nodes, resulting in local amplification and subsequent systemic dissemination by viremia.
Conflict of interest statement
A.D.M.E. Osterhaus wishes to declare, for the avoidance of any misunderstanding on competing interests, that he founded and is chief scientific officer of Viroclinics, a company set up in collaboration with Erasmus University. However, for clarification, no materials or support were received from the company, and no agreements were in place concerning the execution or publication of this work.
Figures
References
-
- Griffin DE. Measles virus. In: Knipe DM, Howley PM, editors. Fields Virology. Philadelphia: Lippincott Williams & Wilkins; 2007. pp. 1551–1585.
-
- WHO. Global reductions in measles mortality 2000–2008 and the risk of measles resurgence. Wkly Epidemiol Rec. 2009;84:509–516. - PubMed
-
- Dörig RE, Marcil A, Chopra A, Richardson CD. The human CD46 molecule is a receptor for measles virus (Edmonston strain). Cell. 1993;75:295–305. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
