

Environmental and genetic preconditioning for long-term anoxia responses requires AMPK in Caenorhabditis elegans
- PMID: 21304820
- PMCID: PMC3033420
- DOI: 10.1371/journal.pone.0016790
Environmental and genetic preconditioning for long-term anoxia responses requires AMPK in Caenorhabditis elegans
Abstract
Background: Preconditioning environments or therapeutics, to suppress the cellular damage associated with severe oxygen deprivation, is of interest to our understanding of diseases associated with oxygen deprivation. Wildtype C. elegans exposed to anoxia enter into a state of suspended animation in which energy-requiring processes reversibly arrest. C. elegans at all developmental stages survive 24-hours of anoxia exposure however, the ability of adult hermaphrodites to survive three days of anoxia significantly decreases. Mutations in the insulin-like signaling receptor (daf-2) and LIN-12/Notch (glp-1) lead to an enhanced long-term anoxia survival phenotype.
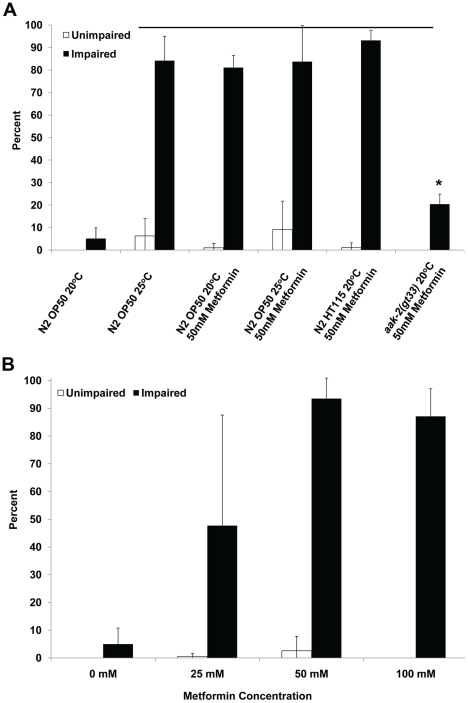
Methodology/principal findings: In this study we show that the combined growth environment of 25°C and a diet of HT115 E. coli will precondition adult hermaphrodites to survive long-term anoxia; many of these survivors have normal movement after anoxia treatment. Animals fed the drug metformin, which induces a dietary-restriction like state in animals and activates AMPK in mammalian cell culture, have a higher survival rate when exposed to long-term anoxia. Mutations in genes encoding components of AMPK (aak-2, aakb-1, aakb-2, aakg-2) suppress the environmentally and genetically induced long-term anoxia survival phenotype. We further determine that there is a correlation between the animals that survive long-term anoxia and increased levels of carminic acid staining, which is a fluorescent dye that incorporates in with carbohydrates such as glycogen.
Conclusions/significance: We conclude that small changes in growth conditions such as increased temperature and food source can influence the physiology of the animal thus affecting the responses to stress such as anoxia. Furthermore, this supports the idea that metformin should be further investigated as a therapeutic tool for treatment of oxygen-deprived tissues. Finally, the capacity for an animal to survive long bouts of severe oxygen deprivation is likely dependent on specific subunits of the heterotrimeric protein AMPK and energy stores such as carbohydrates.
Conflict of interest statement
Figures







Similar articles
-
Reduction in ovulation or male sex phenotype increases long-term anoxia survival in a daf-16-independent manner in Caenorhabditis elegans.Physiol Genomics. 2009 Feb 2;36(3):167-78. doi: 10.1152/physiolgenomics.90278.2008. Epub 2008 Dec 2. Physiol Genomics. 2009. PMID: 19050081 Free PMC article.
-
Glyceraldehyde-3-phosphate dehydrogenase mediates anoxia response and survival in Caenorhabditis elegans.Genetics. 2006 Nov;174(3):1173-87. doi: 10.1534/genetics.106.061390. Epub 2006 Sep 15. Genetics. 2006. PMID: 16980394 Free PMC article.
-
Glucose induces sensitivity to oxygen deprivation and modulates insulin/IGF-1 signaling and lipid biosynthesis in Caenorhabditis elegans.Genetics. 2015 May;200(1):167-84. doi: 10.1534/genetics.115.174631. Epub 2015 Mar 10. Genetics. 2015. PMID: 25762526 Free PMC article.
-
Suspended animation, diapause and quiescence: arresting the cell cycle in C. elegans.Cell Cycle. 2012 May 1;11(9):1672-9. doi: 10.4161/cc.19444. Epub 2012 May 1. Cell Cycle. 2012. PMID: 22510566 Free PMC article. Review.
-
5'-AMP-Activated Protein Kinase Signaling in Caenorhabditis elegans.Exp Suppl. 2016;107:375-388. doi: 10.1007/978-3-319-43589-3_15. Exp Suppl. 2016. PMID: 27812988 Review.
Cited by
-
Effect of dietary nonstructural carbohydrate content on activation of 5'-adenosine monophosphate-activated protein kinase in liver, skeletal muscle, and digital laminae of lean and obese ponies.J Vet Intern Med. 2014 Jul-Aug;28(4):1280-8. doi: 10.1111/jvim.12356. Epub 2014 Apr 20. J Vet Intern Med. 2014. PMID: 24750267 Free PMC article.
-
Anoxia-reoxygenation regulates mitochondrial dynamics through the hypoxia response pathway, SKN-1/Nrf, and stomatin-like protein STL-1/SLP-2.PLoS Genet. 2013;9(12):e1004063. doi: 10.1371/journal.pgen.1004063. Epub 2013 Dec 26. PLoS Genet. 2013. PMID: 24385935 Free PMC article.
-
Identification of enzymes that have helminth-specific active sites and are required for Rhodoquinone-dependent metabolism as targets for new anthelmintics.PLoS Negl Trop Dis. 2021 Nov 29;15(11):e0009991. doi: 10.1371/journal.pntd.0009991. eCollection 2021 Nov. PLoS Negl Trop Dis. 2021. PMID: 34843467 Free PMC article.
-
Repurposing metformin: an old drug with new tricks in its binding pockets.Biochem J. 2015 Nov 1;471(3):307-22. doi: 10.1042/BJ20150497. Biochem J. 2015. PMID: 26475449 Free PMC article. Review.
-
Nicotinamide mononucleotide adenylyltransferase promotes hypoxic survival by activating the mitochondrial unfolded protein response.Cell Death Dis. 2016 Feb 25;7(2):e2113. doi: 10.1038/cddis.2016.5. Cell Death Dis. 2016. PMID: 26913604 Free PMC article.
References
-
- Powers SK, Murlasits Z, Wu M, Kavazis AN. Ischemia-reperfusion-induced cardiac injury: a brief review. Med Sci Sports Exerc. 2007;39:1529–1536. - PubMed
-
- Menger MD, Vollmar B. Pathomechanisms of ischemia-reperfusion injury as the basis for novel preventive strategies: is it time for the introduction of pleiotropic compounds? Transplant Proc. 2007;39:485–488. - PubMed
-
- Pignataro G, Scorziello A, Di Renzo G, Annunziato L. Post-ischemic brain damage: effect of ischemic preconditioning and postconditioning and identification of potential candidates for stroke therapy. FEBS J. 2009;276:46–57. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
