

Stage-specific expression of TNFα regulates bad/bid-mediated apoptosis and RIP1/ROS-mediated secondary necrosis in Birnavirus-infected fish cells
- PMID: 21304825
- PMCID: PMC3033425
- DOI: 10.1371/journal.pone.0016740
Stage-specific expression of TNFα regulates bad/bid-mediated apoptosis and RIP1/ROS-mediated secondary necrosis in Birnavirus-infected fish cells
Abstract
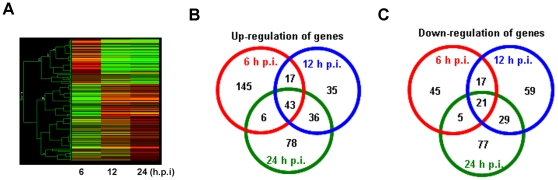
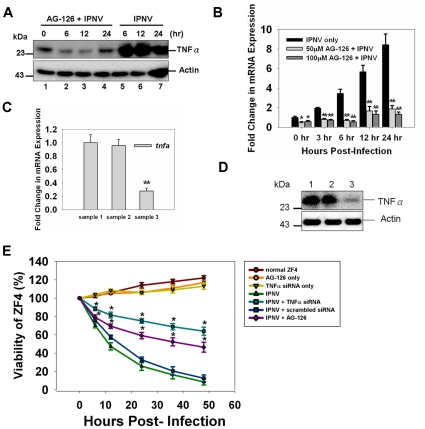
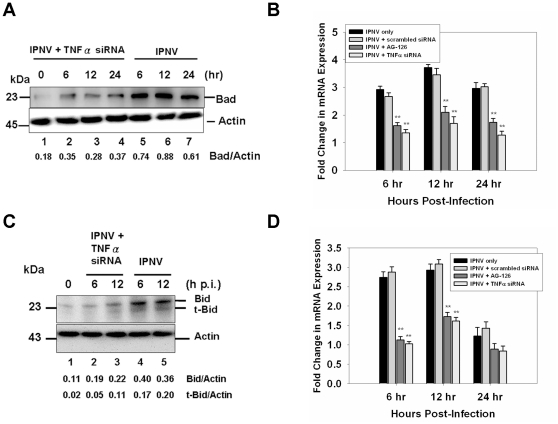
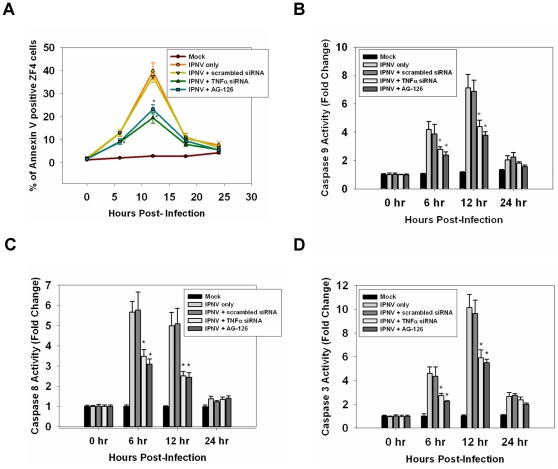
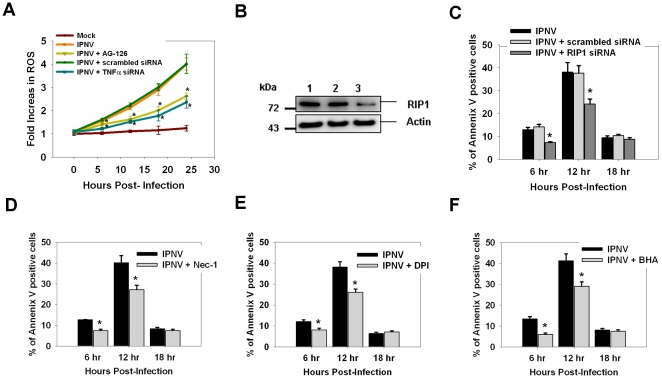
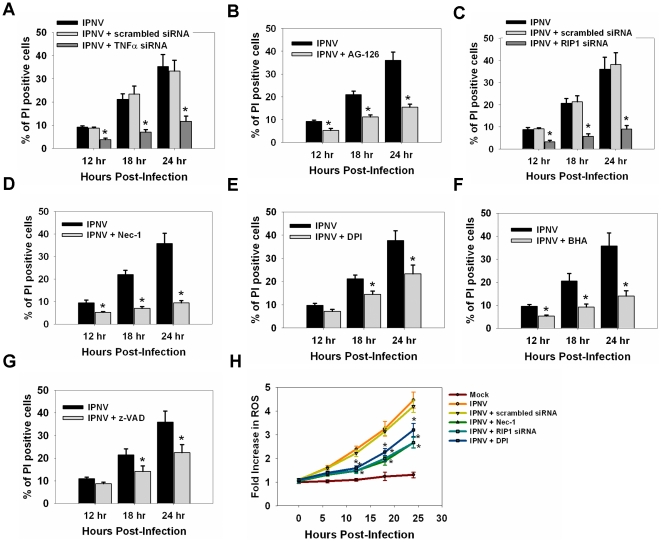
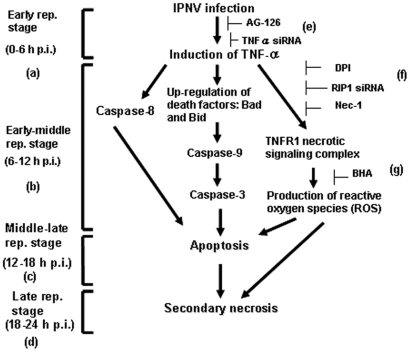
Infectious pancreatic necrosis virus (IPNV) can induce Bad-mediated apoptosis followed by secondary necrosis in fish cells, but it is not known how these two types of cell death are regulated by IPNV. We found that IPNV infection can regulate Bad/Bid-mediated apoptotic and Rip1/ROS-mediated necrotic death pathways via the up-regulation of TNFα in zebrafish ZF4 cells. Using a DNA microarray and quantitative RT-PCR analyses, two major subsets of differentially expressed genes were characterized, including the innate immune response gene TNFα and the pro-apoptotic genes Bad and Bid. In the early replication stage (0-6 h post-infection, or p.i.), we observed that the pro-inflammatory cytokine TNFα underwent a rapid six-fold induction. Then, during the early-middle replication stages (6-12 h p.i.), TNFα level was eight-fold induction and the pro-apoptotic Bcl-2 family members Bad and Bid were up-regulated. Furthermore, specific inhibitors of TNFα expression (AG-126 or TNFα-specific siRNA) were used to block apoptotic and necrotic death signaling during the early or early-middle stages of IPNV infection. Inhibition of TNFα expression dramatically reduced the Bad/Bid-mediated apoptotic and Rip1/ROS-mediated necrotic cell death pathways and rescued host cell viability. Moreover, we used Rip1-specific inhibitors (Nec-1 and Rip1-specific siRNA) to block Rip1 expression. The Rip1/ROS-mediated secondary necrotic pathway appeared to be reduced in IPNV-infected fish cells during the middle-late stage of infection (12-18 h p.i.). Taken together, our results indicate that IPNV triggers two death pathways via up-stream induction of the pro-inflammatory cytokine TNFα, and these results may provide new insights into the pathogenesis of RNA viruses.
Conflict of interest statement
Figures
Similar articles
-
Aquatic birnavirus capsid protein, VP3, induces apoptosis via the Bad-mediated mitochondria pathway in fish and mouse cells.Apoptosis. 2010 Jun;15(6):653-68. doi: 10.1007/s10495-010-0468-x. Apoptosis. 2010. PMID: 20131002
-
Activation of cytokine expression occurs through the TNFα/NF-κB-mediated pathway in birnavirus-infected cells.Fish Shellfish Immunol. 2011 Jul;31(1):10-21. doi: 10.1016/j.fsi.2011.01.015. Epub 2011 Jan 25. Fish Shellfish Immunol. 2011. PMID: 21272652
-
Induction of apoptotic death in cells via Bad gene expression by infectious pancreatic necrosis virus infection.Cell Death Differ. 2002 Feb;9(2):113-24. doi: 10.1038/sj.cdd.4400933. Cell Death Differ. 2002. PMID: 11840162
-
Bid: a Bax-like BH3 protein.Oncogene. 2008 Dec;27 Suppl 1:S93-104. doi: 10.1038/onc.2009.47. Oncogene. 2008. PMID: 19641510 Review.
-
Reactive oxygen species in TNFalpha-induced signaling and cell death.Mol Cells. 2010 Jul;30(1):1-12. doi: 10.1007/s10059-010-0105-0. Epub 2010 Jul 14. Mol Cells. 2010. PMID: 20652490 Free PMC article. Review.
Cited by
-
Pax2 regulates a fadd-dependent molecular switch that drives tissue fusion during eye development.Hum Mol Genet. 2012 May 15;21(10):2357-69. doi: 10.1093/hmg/dds056. Epub 2012 Feb 21. Hum Mol Genet. 2012. PMID: 22357656 Free PMC article.
-
Sulfamethoxazole (SMX) Alters Immune and Apoptotic Endpoints in Developing Zebrafish (Danio rerio).Toxics. 2023 Feb 14;11(2):178. doi: 10.3390/toxics11020178. Toxics. 2023. PMID: 36851053 Free PMC article.
-
Fish TNF and TNF receptors.Sci China Life Sci. 2021 Feb;64(2):196-220. doi: 10.1007/s11427-020-1712-4. Epub 2020 Jul 24. Sci China Life Sci. 2021. PMID: 32720033 Review.
-
Effects of a Dissostichus mawsoni-CaM recombinant proteins feed additive on the juvenile orange-spotted grouper (Epinephelus coioides) under the acute low temperature challenge.Fish Physiol Biochem. 2015 Oct;41(5):1345-58. doi: 10.1007/s10695-015-0090-5. Epub 2015 Jun 30. Fish Physiol Biochem. 2015. PMID: 26122279 Clinical Trial.
-
H(2)O (2)-induced secretion of tumor necrosis factor-α evokes apoptosis of cardiac myocytes through reactive oxygen species-dependent activation of p38 MAPK.Cytotechnology. 2012 Jan;64(1):65-73. doi: 10.1007/s10616-011-9392-3. Epub 2011 Oct 15. Cytotechnology. 2012. PMID: 22002864 Free PMC article.
References
-
- Magyar G, Dobos P. Evidence for the detection of the infectious pancreatic necrosis virus polyprotein and the 17-kDa polypeptide in infected cells and of the NS protease in purified virus. Virology. 1994;204:580–589. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
