

Role of the pJM1 plasmid-encoded transport proteins FatB, C and D in ferric anguibactin uptake in the fish pathogen Vibrio anguillarum
- PMID: 21304833
- PMCID: PMC3034151
- DOI: 10.1111/j.1758-2229.2009.00110.x
Role of the pJM1 plasmid-encoded transport proteins FatB, C and D in ferric anguibactin uptake in the fish pathogen Vibrio anguillarum
Abstract
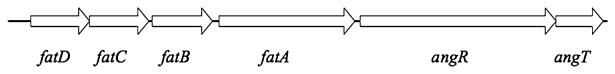
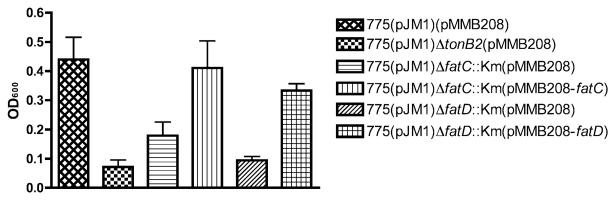
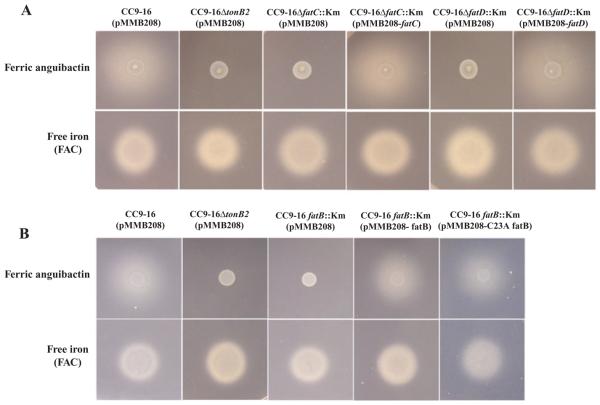
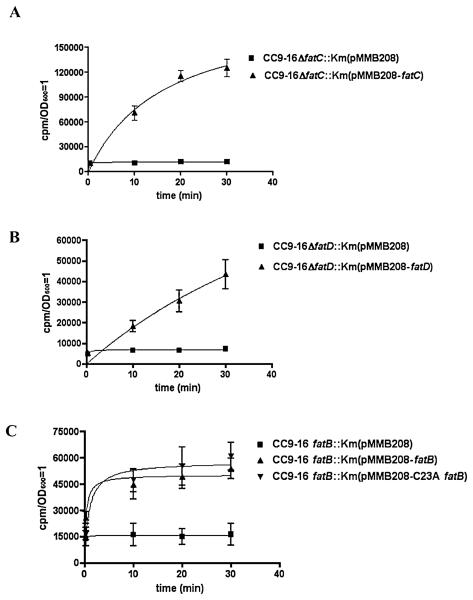
Vibrio anguillarum serotype O1 is part of the natural flora in the aquatic habitat, but under certain circumstances it can cause terminal haemorrhagic septicemia in marine and fresh water fish due to the action of the anguibactin iron uptake system encoded by the virulence plasmid pJM1. This plasmid harbours the genes for the biosynthesis of the siderophore anguibactin and the ferric anguibactin transport proteins FatD, C, B and A encoded in the iron transport operon. The FatA protein is the outer membrane receptor for the ferric siderophore complex and the FatB lipoprotein provides the periplasmic domain for its internalization, whereas the FatC and D proteins are located in the cytoplasmic membrane and might play a role as part of the ABC transporter for internalization of the ferric siderophore. In this work we demonstrate the essential role of these two inner membrane proteins in ferric anguibactin transport and that the lipo-protein nature of FatB is not necessary for ferric anguibactin transport.
Figures
Similar articles
-
Identification of amino acid residues required for ferric-anguibactin transport in the outer-membrane receptor FatA of Vibrio anguillarum.Microbiology (Reading). 2007 Feb;153(Pt 2):570-584. doi: 10.1099/mic.0.2006/001735-0. Microbiology (Reading). 2007. PMID: 17259629
-
Two ABC transporter systems participate in siderophore transport in the marine pathogen Vibrio anguillarum 775 (pJM1).FEMS Microbiol Lett. 2013 Apr;341(2):79-86. doi: 10.1111/1574-6968.12092. Epub 2013 Feb 18. FEMS Microbiol Lett. 2013. PMID: 23350649 Free PMC article.
-
Plasmid-mediated iron uptake and virulence in Vibrio anguillarum.Plasmid. 2002 Nov;48(3):222-8. doi: 10.1016/s0147-619x(02)00111-7. Plasmid. 2002. PMID: 12460538
-
Characterization of ferric-anguibactin transport in Vibrio anguillarum.Biometals. 2007 Jun;20(3-4):393-403. doi: 10.1007/s10534-007-9084-9. Epub 2007 Feb 8. Biometals. 2007. PMID: 17287889 Review.
-
Regulation of plasmid-mediated iron transport and virulence in Vibrio anguillarum.Biol Met. 1991;4(1):33-5. doi: 10.1007/BF01135554. Biol Met. 1991. PMID: 1854586 Review.
Cited by
-
Fitness factors in vibrios: a mini-review.Microb Ecol. 2013 May;65(4):826-51. doi: 10.1007/s00248-012-0168-x. Epub 2013 Jan 10. Microb Ecol. 2013. PMID: 23306394 Review.
-
Modulation of Bacterial Fitness and Virulence Through Antisense RNAs.Front Cell Infect Microbiol. 2021 Feb 11;10:596277. doi: 10.3389/fcimb.2020.596277. eCollection 2020. Front Cell Infect Microbiol. 2021. PMID: 33747974 Free PMC article. Review.
-
The influence of the accessory genome on bacterial pathogen evolution.Mob Genet Elements. 2011 May;1(1):55-65. doi: 10.4161/mge.1.1.16432. Mob Genet Elements. 2011. PMID: 22016845 Free PMC article.
-
Genetic Determinants of Virulence in the Marine Fish Pathogen Vibrio anguillarum.Fish Pathol. 2011 Jan 1;46:1-10. doi: 10.3147/jsfp.46.1. Fish Pathol. 2011. PMID: 21625345 Free PMC article.
-
Iron Acquisition Strategies of Vibrio anguillarum.Front Cell Infect Microbiol. 2017 Jul 25;7:342. doi: 10.3389/fcimb.2017.00342. eCollection 2017. Front Cell Infect Microbiol. 2017. PMID: 28791260 Free PMC article. Review.
References
-
- Actis LA, Tolmasky ME, Farrell DH, Crosa JH. Genetic and molecular characterization of essential components of the Vibrio anguillarum plasmid-mediated iron-transport system. J Biol Chem. 1988;263:2853–2860. - PubMed
-
- Actis LA, Tolmasky ME, Crosa LM, Crosa JH. Characterization and regulation of the expression of FatB, an iron transport protein encoded by the pJM1 virulence plasmid. Mol Microbiol. 1995;17:197–204. - PubMed
-
- Actis LA, Tolmasky ME, Crosa JH. Vibriosis. Vol. 3. Cab International Publishing; Wallingford, UK: 1999.
Grants and funding
LinkOut - more resources
Full Text Sources
