

Signaling pathways that control cell migration: models and analysis
- PMID: 21305705
- PMCID: PMC3052860
- DOI: 10.1002/wsbm.110
Signaling pathways that control cell migration: models and analysis
Abstract
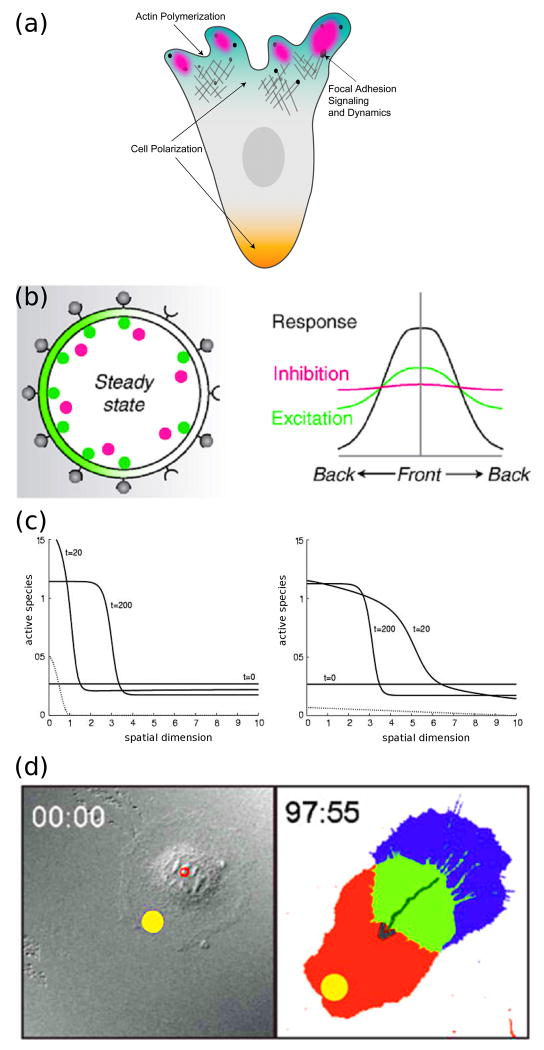
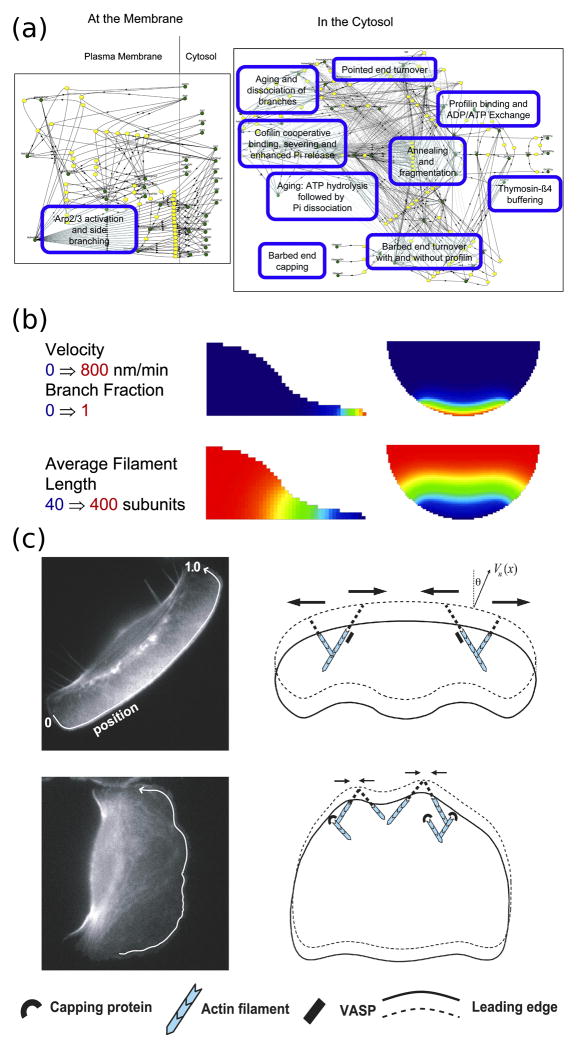
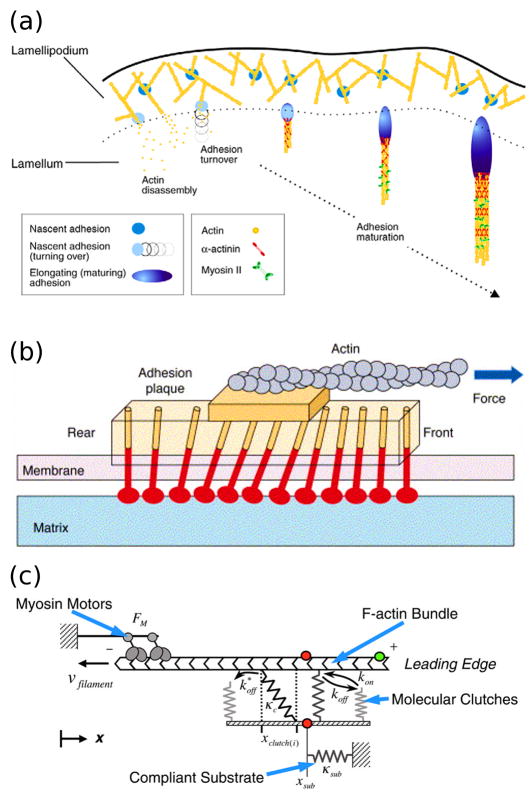
Dissecting the intracellular signaling mechanisms that govern the movement of eukaryotic cells presents a major challenge, not only because of the large number of molecular players involved, but even more so because of the dynamic nature of their regulation by both biochemical and mechanical interactions. Computational modeling and analysis have emerged as useful tools for understanding how the physical properties of cells and their microenvironment are coupled with certain biochemical pathways to actuate and control cell motility. In this focused review, we highlight some of the more recent applications of quantitative modeling and analysis in the field of cell migration. Both in modeling and experiment, it has been prudent to follow a reductionist approach in order to characterize what are arguably the principal modules: spatial polarization of signaling pathways, regulation of the actin cytoskeleton, and dynamics of focal adhesions. While it is important that we 'cut our teeth' on these subsystems, focusing on the details of certain aspects while ignoring or coarse-graining others, it is clear that the challenge ahead will be to characterize the couplings between them in an integrated framework.
Copyright © 2010 John Wiley & Sons, Inc.
Figures
References
-
- Ridley AJ, Schwartz MA, Burridge K, Firtel RA, Ginsberg MH, Borisy G, Parsons TJ, Horwitz AR. Cell migration: Integrating signals from front to back. Science. 2003;302:1704–1709. - PubMed
Further Reading
-
- Pollard TD, Borisy GG. Cellular motility driven by assembly and disassembly of actin filaments. Cell. 2003;112:453–465. - PubMed
-
- Dorn JF, Danuser G, Yang G. Computational processing and analysis of dynamic fluorescence image data. Methods Cell Biol. 2008;85:497–538. - PubMed
-
- Wang Y. Flux at focal adhesions: Slippage clutch, mechanical gauge, or signal depot. Sci STKE. 2007;377:pe10. - PubMed
-
- Orr AW, Helmke BP, Blackman BR, Schwartz MA. Mechanisms of mechanotransduction. Dev Cell. 2006;10:11–20. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
