

Pollen tube growth and guidance: roles of small, secreted proteins
- PMID: 21307038
- PMCID: PMC3170145
- DOI: 10.1093/aob/mcr015
Pollen tube growth and guidance: roles of small, secreted proteins
Abstract
Background: Pollination is a crucial step in angiosperm (flowering plant) reproduction. Highly orchestrated pollen-pistil interactions and signalling events enable plant species to avoid inbreeding and outcrossing as a species-specific barrier. In compatible pollination, pollen tubes carrying two sperm cells grow through the pistil transmitting tract and are precisely guided to the ovules, discharging the sperm cells to the embryo sac for fertilization.
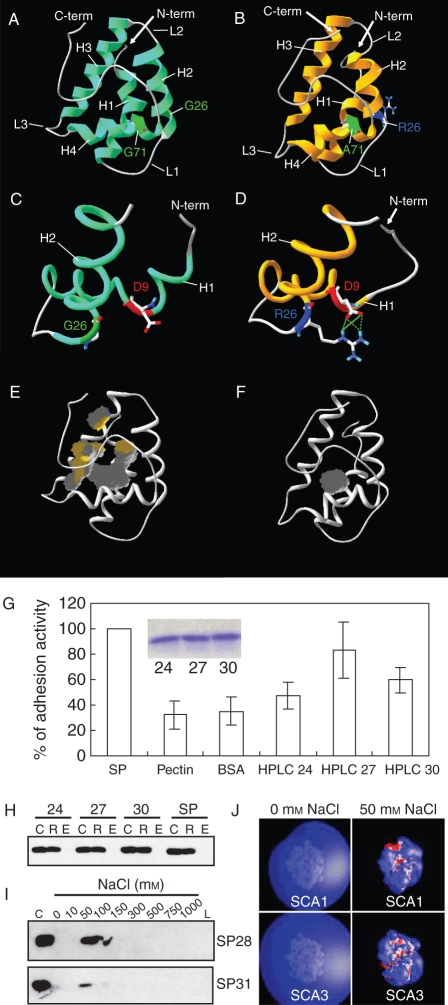
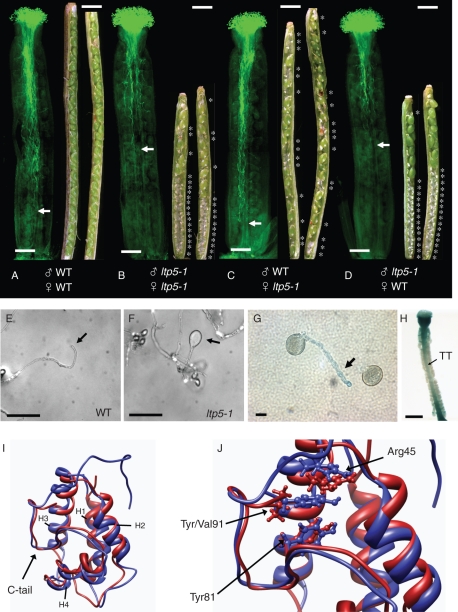
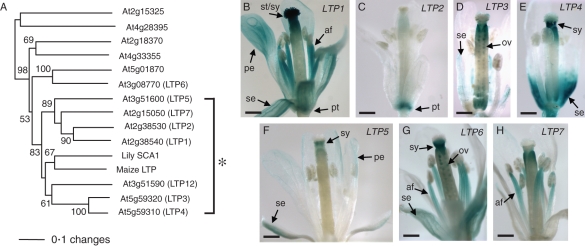
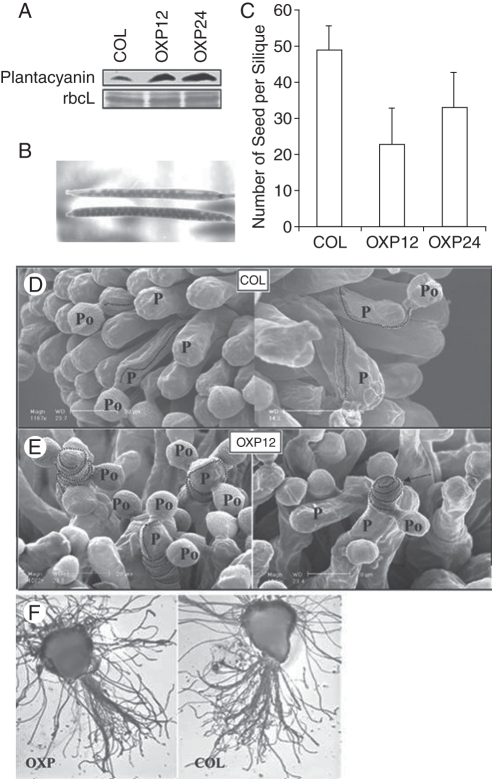
Scope: In Lilium longiflorum pollination, growing pollen tubes utilize two critical mechanisms, adhesion and chemotropism, for directional growth to the ovules. Among several molecular factors discovered in the past decade, two small, secreted cysteine-rich proteins have been shown to play major roles in pollen tube adhesion and reorientation bioassays: stigma/style cysteine-rich adhesin (SCA, approx. 9·3 kDa) and chemocyanin (approx. 9·8 kDa). SCA, a lipid transfer protein (LTP) secreted from the stylar transmitting tract epidermis, functions in lily pollen tube tip growth as well as in forming the adhesive pectin matrix at the growing pollen tube wall back from the tip. Lily chemocyanin is a plantacyanin family member and acts as a directional cue for reorienting pollen tubes. Recent consecutive studies revealed that Arabidopsis thaliana homologues for SCA and chemocyanin play pivotal roles in tip polarity and directionality of pollen tube growth, respectively. This review outlines the biological roles of various secreted proteins in angiosperm pollination, focusing on plant LTPs and chemocyanin.
Figures
References
-
- Arondel V, Vergnolle C, Cantrel C, Kader JC. Lipid transfer proteins are encoded by a small multigene family in Arabidopsis thaliana. Plant Science. 2000;157:1–12. - PubMed
-
- von Besser K, Frank AC, Johnson MA, Preuss D. Arabidopsis HAP2 (GCS1) is a sperm-specific gene required for pollen tube guidance and fertilization. Development. 2006;133:4761–4769. - PubMed
-
- Bosch M, Franklin-Tong VE. Self-incompatibility in Papaver: signalling to trigger PCD in incompatible pollen. Journal of Experimental Botany. 2008;59:481–490. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
