

The IRF-3/Bax-mediated apoptotic pathway, activated by viral cytoplasmic RNA and DNA, inhibits virus replication
- PMID: 21307205
- PMCID: PMC3126131
- DOI: 10.1128/JVI.02133-10
The IRF-3/Bax-mediated apoptotic pathway, activated by viral cytoplasmic RNA and DNA, inhibits virus replication
Abstract
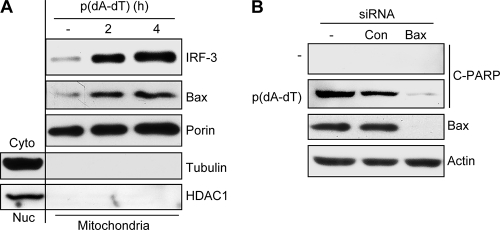
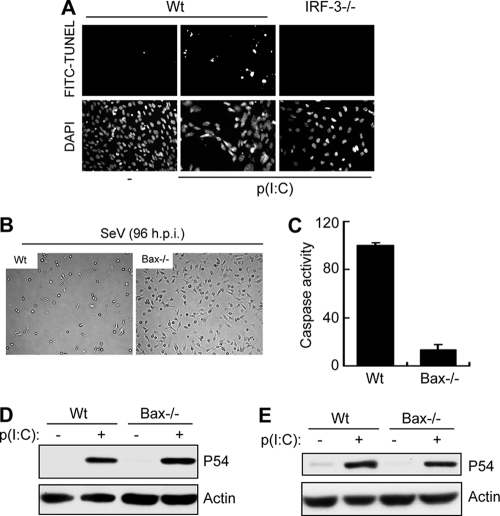
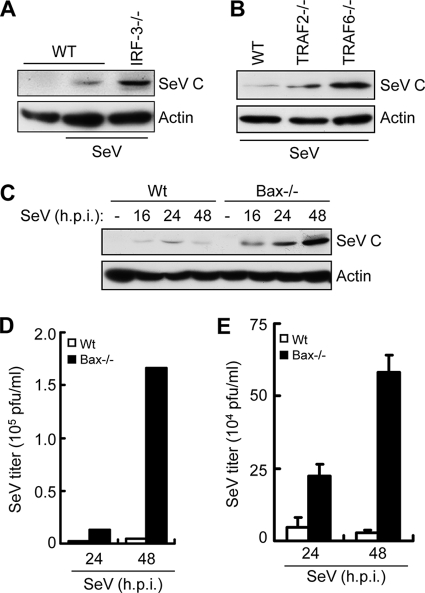
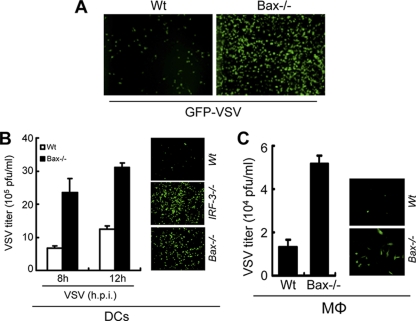
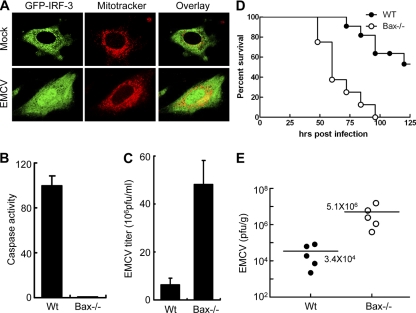
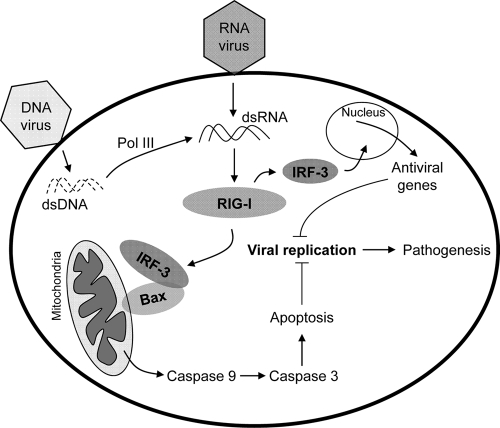
Induction of apoptosis in cells infected by Sendai virus (SeV), which triggers the cytosolic RIG-I pathway, requires the presence of interferon regulatory factor 3 (IRF-3). Independent of IRF-3's transcriptional role, a novel IRF-3 activation pathway causes its interaction with the proapoptotic protein Bax and its mitochondrial translocation to induce apoptosis. Here we report that two other RNA viruses, vesicular stomatitis virus (VSV) and encephalomyocarditis virus (EMCV), may also activate the same pathway. Moreover, cytosolic DNA, produced by adenovirus or introduced by transfection, activated the pathway in an RNA polymerase III-dependent fashion. To evaluate the contribution of this newly discovered apoptotic pathway to the host's overall antiviral response, we measured the efficiencies of replication of various viruses in vitro and viral pathogenesis in vivo, using cells and mice that are selectively deficient in components required for the apoptotic pathway of IRF-3. Our results clearly demonstrate that the IRF-3/Bax-mediated apoptotic signaling branch contributes significantly to the host's protection from viral infection and consequent pathogenesis.
Figures
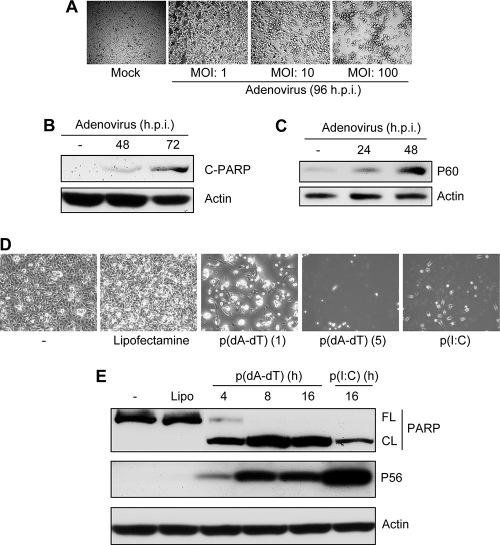
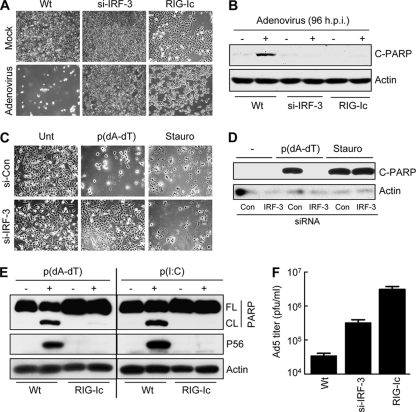
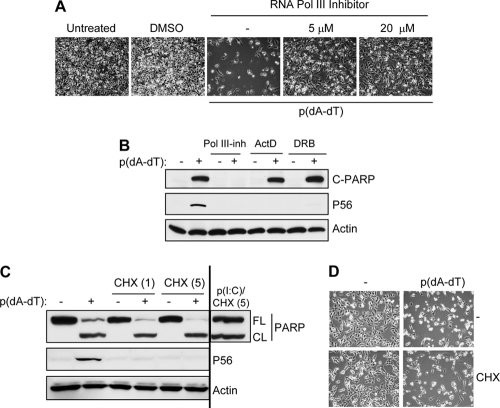
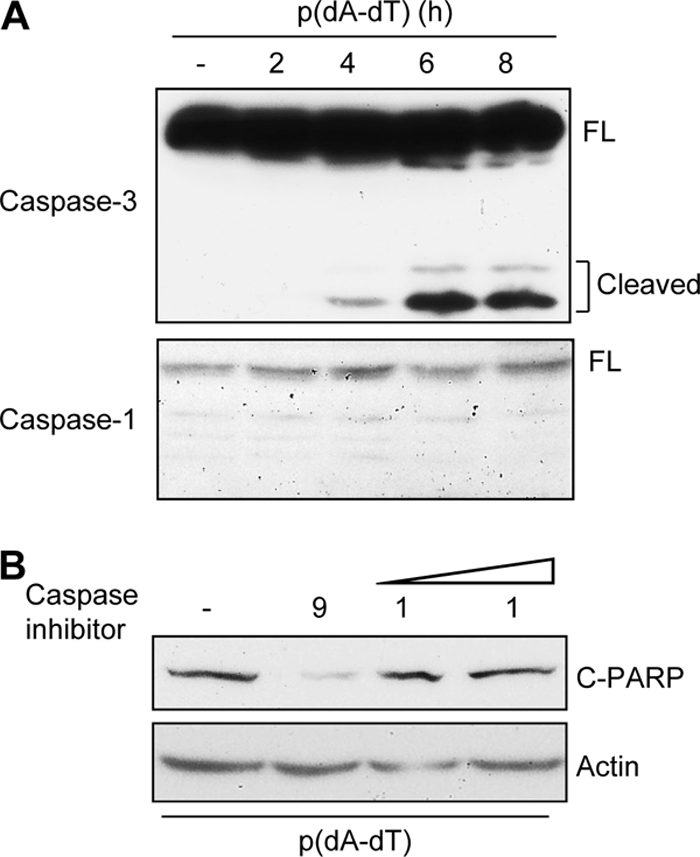
References
-
- Alexopoulou L., Holt A. C., Medzhitov R., Flavell R. A. 2001. Recognition of double-stranded RNA and activation of NF-kappaB by Toll-like receptor 3. Nature 413:732–738 - PubMed
-
- Andersen J., VanScoy S., Cheng T. F., Gomez D., Reich N. C. 2008. IRF-3-dependent and augmented target genes during viral infection. Genes Immun. 9:168–175 - PubMed
-
- Burckstummer T., et al. 2009. An orthogonal proteomic-genomic screen identifies AIM2 as a cytoplasmic DNA sensor for the inflammasome. Nat. Immunol. 10:266–272 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
