

LRRK2 controls synaptic vesicle storage and mobilization within the recycling pool
- PMID: 21307259
- PMCID: PMC6633036
- DOI: 10.1523/JNEUROSCI.3730-10.2011
LRRK2 controls synaptic vesicle storage and mobilization within the recycling pool
Abstract
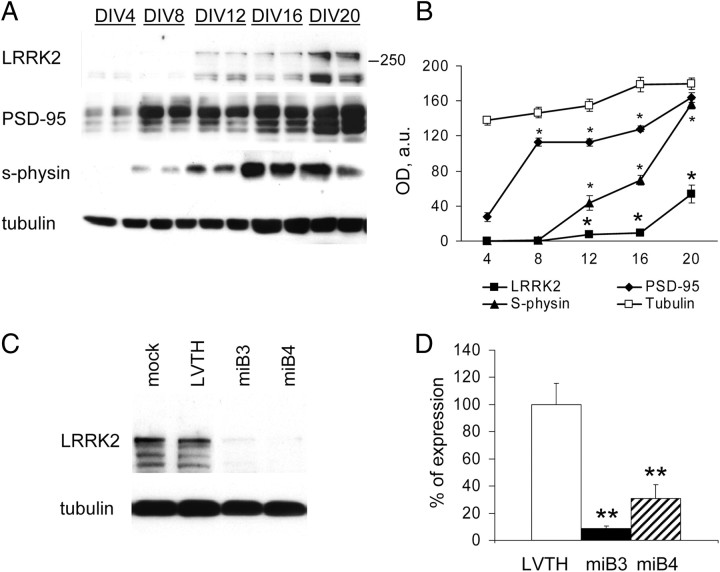
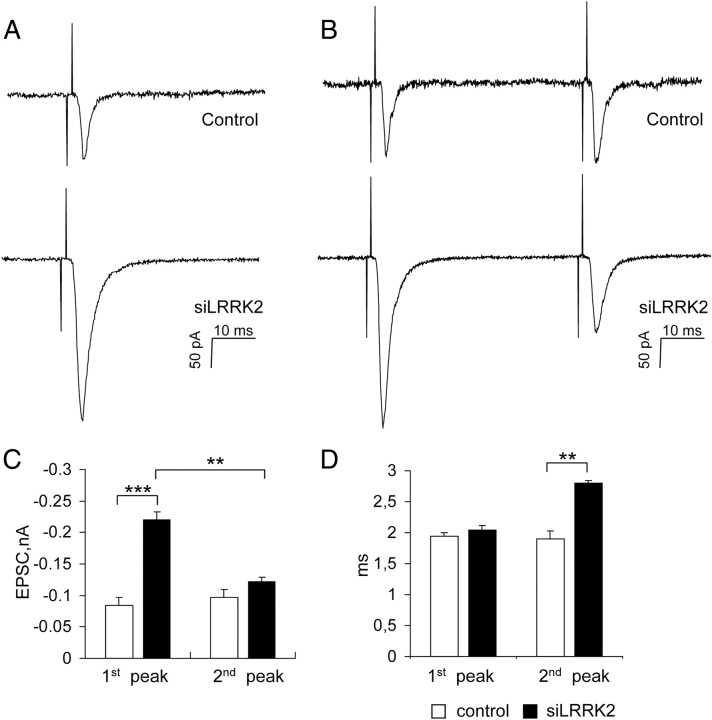
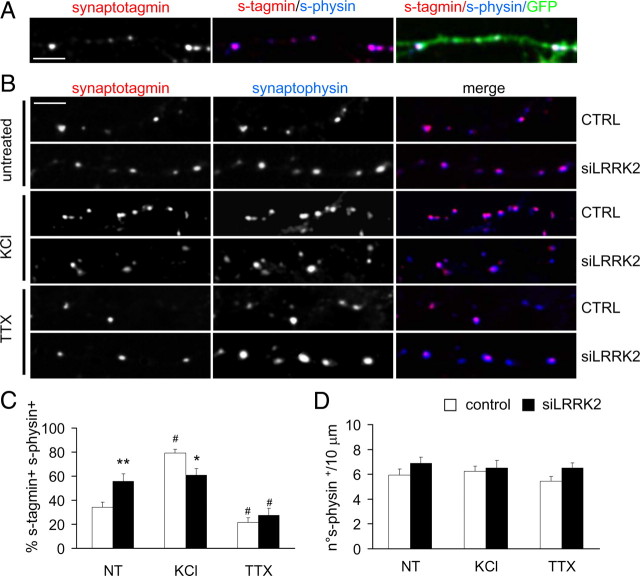
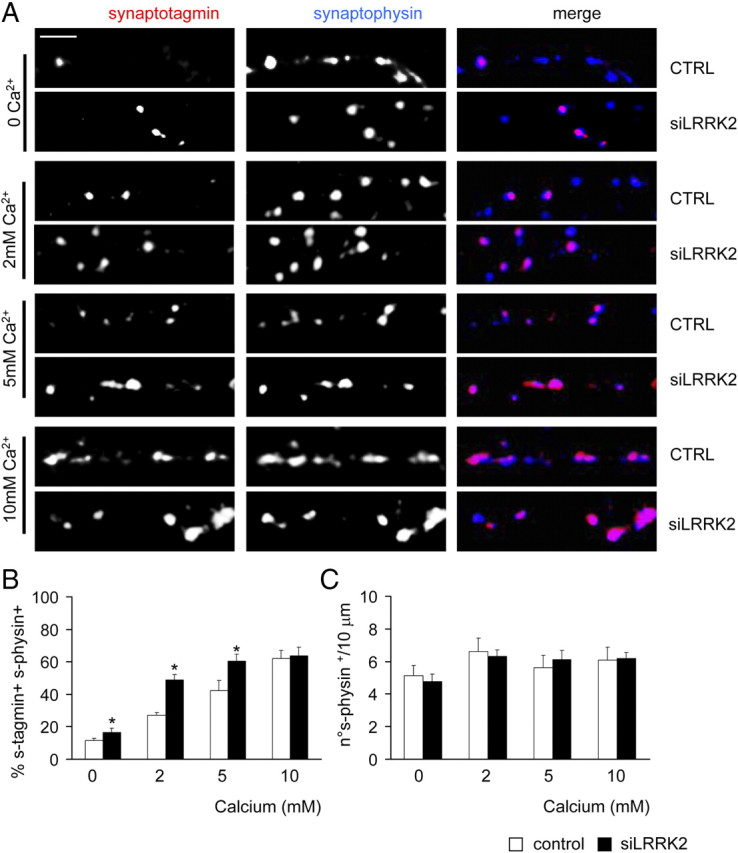
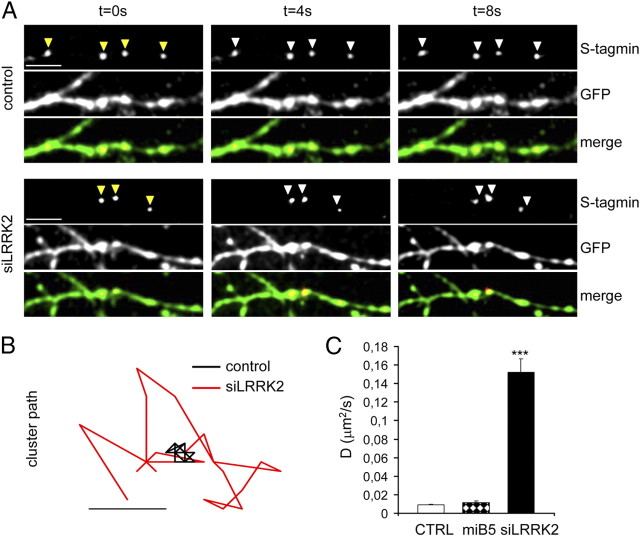
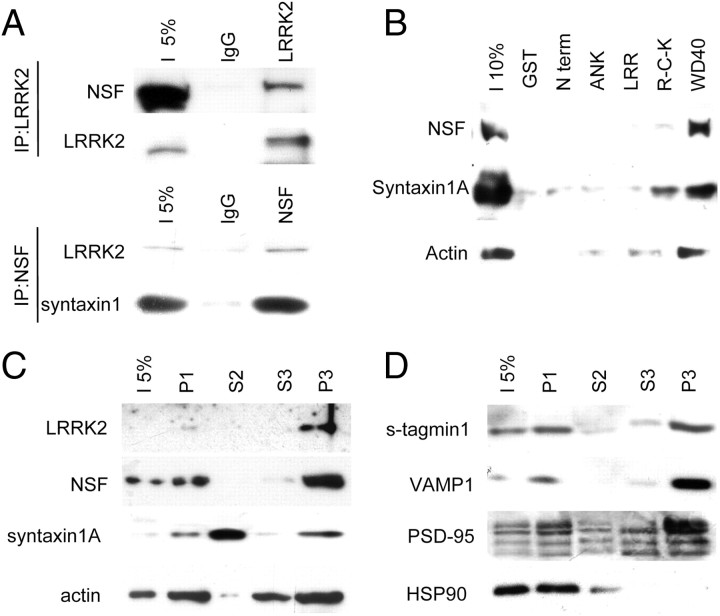
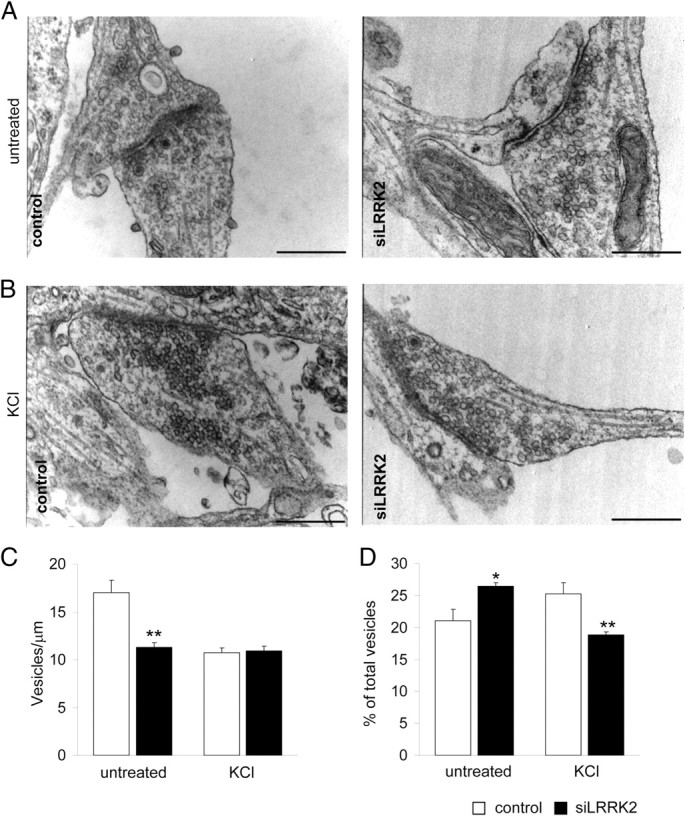
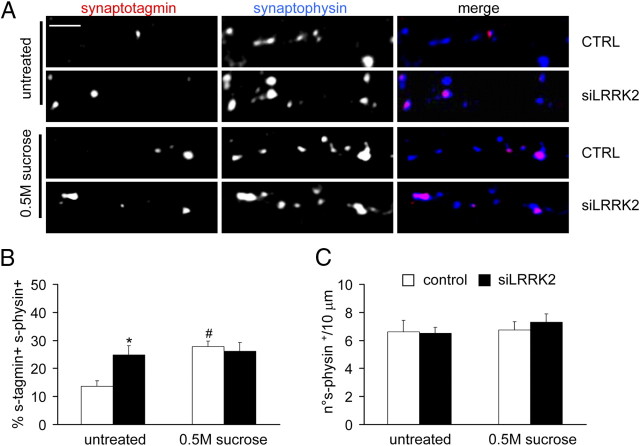
Mutations in leucine-rich repeat kinase 2 (LRRK2) are the single most common cause of inherited Parkinson's disease. Little is known about its involvement in the pathogenesis of Parkinson's disease mainly because of the lack of knowledge about the physiological role of LRRK2. To determine the function of LRRK2, we studied the impact of short hairpin RNA-mediated silencing of LRRK2 expression in cortical neurons. Paired recording indicated that LRRK2 silencing affects evoked postsynaptic currents. Furthermore, LRRK2 silencing induces at the presynaptic site a redistribution of vesicles within the bouton, altered recycling dynamics, and increased vesicle kinetics. Accordingly, LRRK2 protein is present in the synaptosomal compartment of cortical neurons in which it interacts with several proteins involved in vesicular recycling. Our results suggest that LRRK2 modulates synaptic vesicle trafficking and distribution in neurons and in consequence participates in regulating the dynamics between vesicle pools inside the presynaptic bouton.
Figures
Comment in
-
LRRK2: a problem lurking in vesicle trafficking?J Neurosci. 2011 Jul 6;31(27):9787-8. doi: 10.1523/JNEUROSCI.1976-11.2011. J Neurosci. 2011. PMID: 21734269 Free PMC article. No abstract available.
References
-
- Aasly JO, Toft M, Fernandez-Mata I, Kachergus J, Hulihan M, White LR, Farrer M. Clinical features of LRRK2-associated Parkinson's disease in central Norway. Ann Neurol. 2005;57:762–765. - PubMed
-
- Andres-Mateos E, Mejias R, Sasaki M, Li X, Lin BM, Biskup S, Zhang L, Banerjee R, Thomas B, Yang L, Liu G, Beal MF, Huso DL, Dawson TM, Dawson VL. Unexpected lack of hypersensitivity in LRRK2 knock-out mice to MPTP (1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine) J Neurosci. 2009;29:15846–15850. - PMC - PubMed
-
- Augustin I, Rosenmund C, Südhof TC, Brose N. Munc13-1 is essential for fusion competence of glutamatergic synaptic vesicles. Nature. 1999;400:457–461. - PubMed
-
- Bacci A, Coco S, Pravettoni E, Schenk U, Armano S, Frassoni C, Verderio C, De Camilli P, Matteoli M. Chronic blockade of glutamate receptors enhances presynaptic release and downregulates the interaction between synaptophysin-synaptobrevin-vesicle-associated membrane protein 2. J Neurosci. 2001;21:6588–6596. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases