

Regulation of MiR-124, Let-7d, and MiR-181a in the accumbens affects the expression, extinction, and reinstatement of cocaine-induced conditioned place preference
- PMID: 21307844
- PMCID: PMC3079833
- DOI: 10.1038/npp.2010.250
Regulation of MiR-124, Let-7d, and MiR-181a in the accumbens affects the expression, extinction, and reinstatement of cocaine-induced conditioned place preference
Abstract
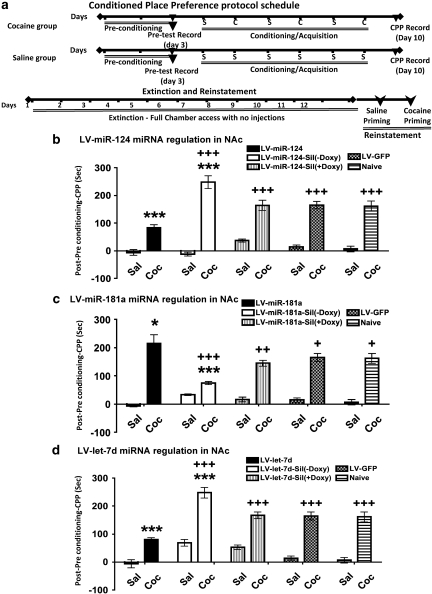
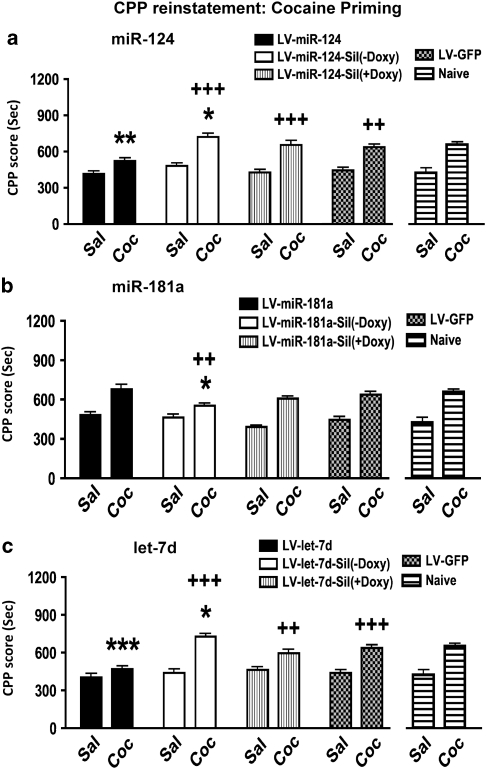
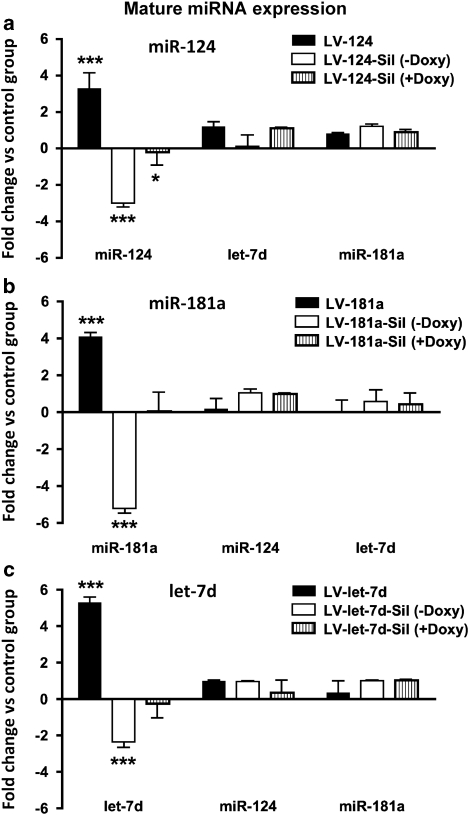
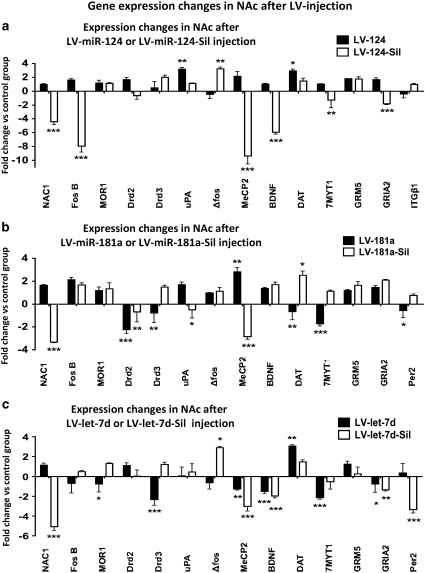
Molecular adaptations underlying drug seeking and relapse remain largely unknown. Studies highlight post-transcriptional modifications mediated by microRNAs (miRNAs) in addiction and other neurological disorders. We have previously shown that chronic cocaine suppresses miR-124 and let-7d and induces the expression of miR-181a in mesolimbic pathway. To further address the role and target gene regulation network of these miRNAs in vivo in cocaine addiction, we developed lentiviral vector (LV)-expressing miRNAs and their corresponding silencers for stable and regulatable miRNA expression. We tested in vivo miRNA gain and loss of function on cocaine-induced conditioned place preference (CPP) by localized LV-miRNA regulation in the nucleus accumbens (NAc). LV-miR-124 and let-7d expression in the NAc attenuates cocaine CPP, whereas LV-miR-181a enhances it. Silencing miRNAs by corresponding LV-miRNA silencers expressing perfect miRNA target sequences inversed this effect on cocaine CPP. Doxycycline treatment for switching off silencer expression abolished the observed behavioral changes. Behavioral changes mediated by LV-miRNA regulation resulted in dynamic alterations in transcription factors, receptors, and other effector genes involved in cocaine-induced plasticity. Our results describe a complex regulatory pathway mediated by miRNAs in cocaine-mediated neuronal adaptations.
Figures



Similar articles
-
Role of accumbens BDNF and TrkB in cocaine-induced psychomotor sensitization, conditioned-place preference, and reinstatement in rats.Psychopharmacology (Berl). 2008 Aug;199(2):169-82. doi: 10.1007/s00213-008-1164-1. Epub 2008 Jun 13. Psychopharmacology (Berl). 2008. PMID: 18551281
-
microRNAs miR-124, let-7d and miR-181a regulate cocaine-induced plasticity.Mol Cell Neurosci. 2009 Dec;42(4):350-62. doi: 10.1016/j.mcn.2009.08.009. Epub 2009 Aug 22. Mol Cell Neurosci. 2009. PMID: 19703567
-
In silico identification and in vivo validation of miR-495 as a novel regulator of motivation for cocaine that targets multiple addiction-related networks in the nucleus accumbens.Mol Psychiatry. 2018 Feb;23(2):434-443. doi: 10.1038/mp.2016.238. Epub 2017 Jan 3. Mol Psychiatry. 2018. PMID: 28044061 Free PMC article.
-
Extinction training regulates neuroadaptive responses to withdrawal from chronic cocaine self-administration.Learn Mem. 2004 Sep-Oct;11(5):648-57. doi: 10.1101/lm.81404. Learn Mem. 2004. PMID: 15466321 Free PMC article. Review.
-
Molecular, cellular, and structural mechanisms of cocaine addiction: a key role for microRNAs.Neuropsychopharmacology. 2013 Jan;38(1):198-211. doi: 10.1038/npp.2012.120. Epub 2012 Sep 12. Neuropsychopharmacology. 2013. PMID: 22968819 Free PMC article. Review.
Cited by
-
microRNA regulation related to the protective effects of environmental enrichment against cocaine-seeking behavior.Drug Alcohol Depend. 2021 Apr 1;221:108585. doi: 10.1016/j.drugalcdep.2021.108585. Epub 2021 Feb 16. Drug Alcohol Depend. 2021. PMID: 33647589 Free PMC article.
-
Increased expression of plasma hsa-miR-181a in male patients with heroin addiction use disorder.J Clin Lab Anal. 2020 Nov;34(11):e23486. doi: 10.1002/jcla.23486. Epub 2020 Aug 3. J Clin Lab Anal. 2020. PMID: 32748469 Free PMC article.
-
Peripheral blood microRNA levels in females with cocaine use disorder.J Psychiatr Res. 2019 Jul;114:48-54. doi: 10.1016/j.jpsychires.2019.03.028. Epub 2019 Apr 2. J Psychiatr Res. 2019. PMID: 31026664 Free PMC article.
-
MicroRNA-423-5p Mediates Cocaine-Induced Smooth Muscle Cell Contraction by Targeting Cacna2d2.Int J Mol Sci. 2023 Apr 1;24(7):6584. doi: 10.3390/ijms24076584. Int J Mol Sci. 2023. PMID: 37047559 Free PMC article.
-
MicroRNA expression signature of methamphetamine use and addiction in the rat nucleus accumbens.Metab Brain Dis. 2017 Dec;32(6):1767-1783. doi: 10.1007/s11011-017-0061-x. Epub 2017 Jul 5. Metab Brain Dis. 2017. PMID: 28681200
References
-
- Ambros V. The functions of animal microRNAs. Nature. 2004;431:350–355. - PubMed
-
- Artinian J, De Jaeger X, Fellini L, de Saint Blanquat P, Roullet P. Reactivation with a simple exposure to the experimental environment is sufficient to induce reconsolidation requiring protein synthesis in the hippocampal CA3 region in mice. Hippocampus. 2007;17:181–191. - PubMed
-
- Ashraf SI, McLoon AL, Sclarsic SM, Kunes S. Synaptic protein synthesis associated with memory is regulated by the RISC pathway in Drosophila. Cell. 2006;124:191–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
