

Crystal structure of the sensory domain of Escherichia coli CadC, a member of the ToxR-like protein family
- PMID: 21308846
- PMCID: PMC3081544
- DOI: 10.1002/pro.594
Crystal structure of the sensory domain of Escherichia coli CadC, a member of the ToxR-like protein family
Abstract
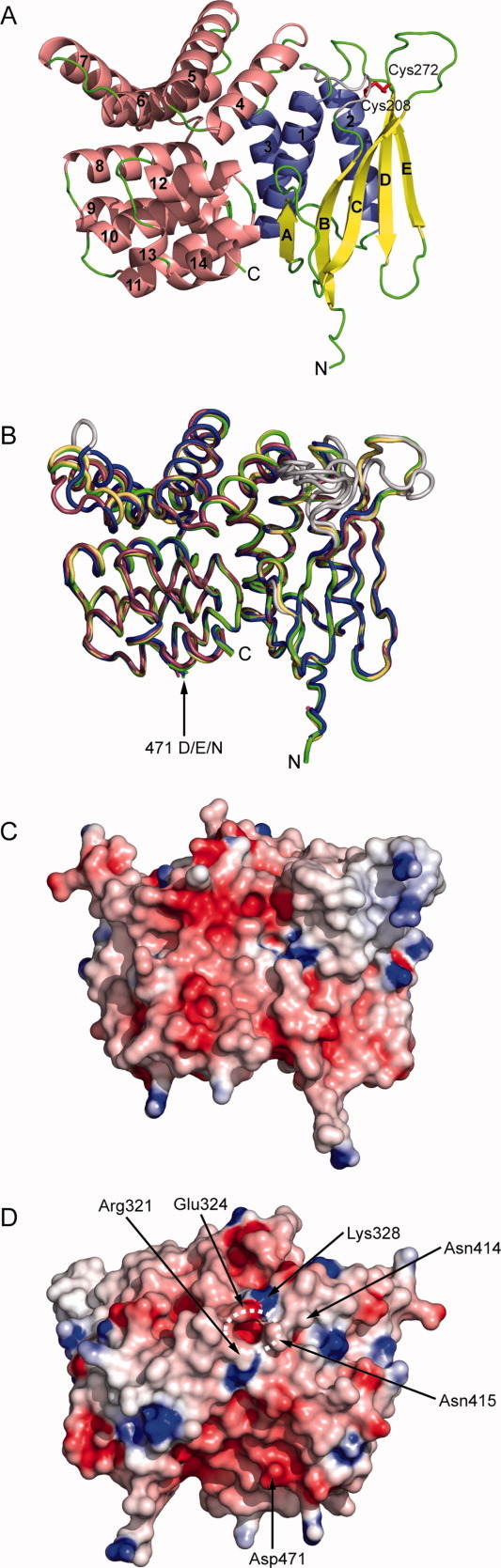
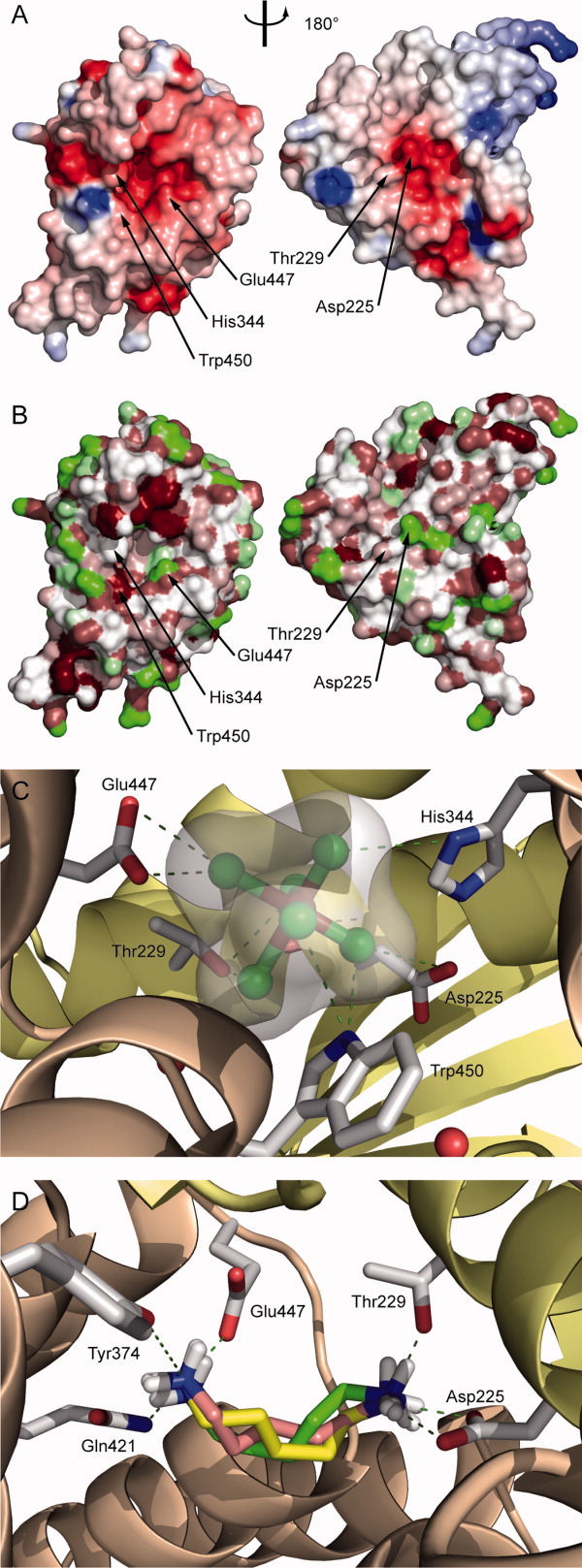
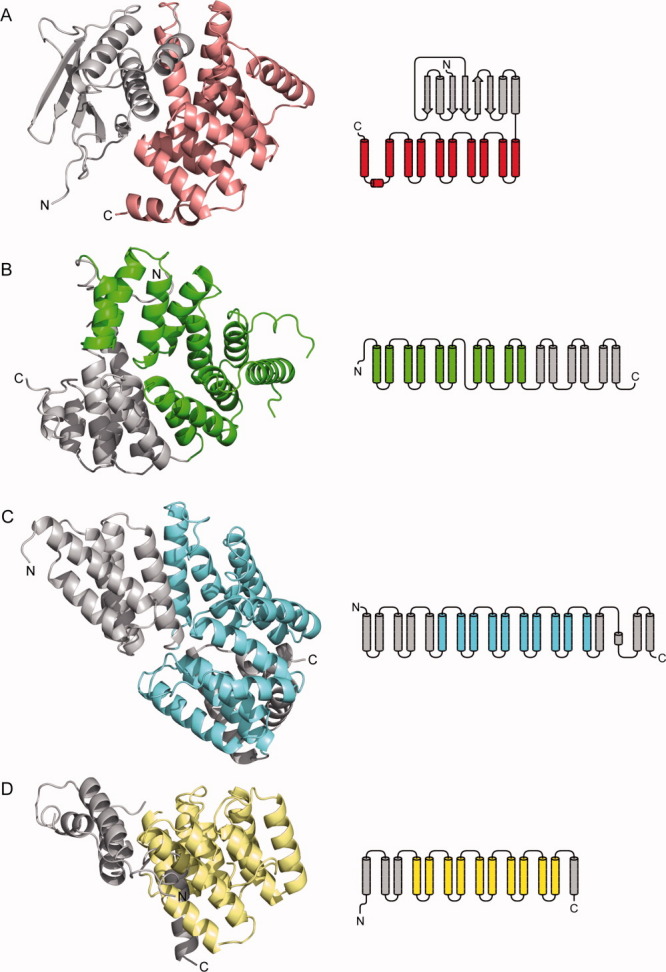
The membrane-integral transcriptional activator CadC comprises sensory and transcriptional regulatory functions within one polypeptide chain. Its C-terminal periplasmic domain, CadC(pd), is responsible for sensing of environmental pH as well as for binding of the feedback inhibitor cadaverine. Here we describe the crystal structure of CadC(pd) (residues 188-512) solved at a resolution of 1.8 Å via multiple wavelength anomalous dispersion (MAD) using a ReCl(6)(2-) derivative. CadC(pd) reveals a novel fold comprising two subdomains: an N-terminal subdomain dominated by a β-sheet in contact with three α-helices and a C-terminal subdomain formed by an eleven-membered α-helical bundle, which is oriented almost perpendicular to the helices in the first subdomain. Further to the native protein, crystal structures were also solved for its variants D471N and D471E, which show functionally different behavior in pH sensing. Interestingly, in the heavy metal derivative of CadC(pd) used for MAD phasing a ReCl(6)(2-) ion was found in a cavity located between the two subdomains. Amino acid side chains that coordinate this complex ion are conserved in CadC homologues from various bacterial species, suggesting a function of the cavity in the binding of cadaverine, which was supported by docking studies. Notably, CadC(pd) forms a homo-dimer in solution, which can be explained by an extended, albeit rather polar interface between two symmetry-related monomers in the crystal structure. The occurrence of several acidic residues in this region suggests protonation-dependent changes in the mode of dimerization, which could eventually trigger transcriptional activation by CadC in the bacterial cytoplasm.
Copyright © 2011 The Protein Society.
Figures


Similar articles
-
New insights into the signaling mechanism of the pH-responsive, membrane-integrated transcriptional activator CadC of Escherichia coli.J Biol Chem. 2011 Mar 25;286(12):10681-9. doi: 10.1074/jbc.M110.196923. Epub 2011 Jan 7. J Biol Chem. 2011. PMID: 21216950 Free PMC article.
-
Detection and function of an intramolecular disulfide bond in the pH-responsive CadC of Escherichia coli.BMC Microbiol. 2011 Apr 12;11:74. doi: 10.1186/1471-2180-11-74. BMC Microbiol. 2011. PMID: 21486484 Free PMC article.
-
Structural and Functional Analysis of the Signal-Transducing Linker in the pH-Responsive One-Component System CadC of Escherichia coli.J Mol Biol. 2015 Jul 31;427(15):2548-2561. doi: 10.1016/j.jmb.2015.05.001. Epub 2015 May 12. J Mol Biol. 2015. PMID: 25979249
-
Structure-function analysis of the DNA-binding domain of a transmembrane transcriptional activator.Sci Rep. 2017 Apr 21;7(1):1051. doi: 10.1038/s41598-017-01031-9. Sci Rep. 2017. PMID: 28432336 Free PMC article.
-
Deactivation of the E. coli pH stress sensor CadC by cadaverine.J Mol Biol. 2012 Nov 23;424(1-2):15-27. doi: 10.1016/j.jmb.2012.08.023. Epub 2012 Sep 18. J Mol Biol. 2012. PMID: 22999955
Cited by
-
New insights into the signaling mechanism of the pH-responsive, membrane-integrated transcriptional activator CadC of Escherichia coli.J Biol Chem. 2011 Mar 25;286(12):10681-9. doi: 10.1074/jbc.M110.196923. Epub 2011 Jan 7. J Biol Chem. 2011. PMID: 21216950 Free PMC article.
-
Structural insights into RNA polymerase recognition and essential function of Myxococcus xanthus CdnL.PLoS One. 2014 Oct 1;9(10):e108946. doi: 10.1371/journal.pone.0108946. eCollection 2014. PLoS One. 2014. PMID: 25272012 Free PMC article.
-
Co-component signal transduction systems: Fast-evolving virulence regulation cassettes discovered in enteric bacteria.Proc Natl Acad Sci U S A. 2022 Jun 14;119(24):e2203176119. doi: 10.1073/pnas.2203176119. Epub 2022 Jun 1. Proc Natl Acad Sci U S A. 2022. PMID: 35648808 Free PMC article.
-
Detection and function of an intramolecular disulfide bond in the pH-responsive CadC of Escherichia coli.BMC Microbiol. 2011 Apr 12;11:74. doi: 10.1186/1471-2180-11-74. BMC Microbiol. 2011. PMID: 21486484 Free PMC article.
-
Bacterial moving and shaking: the 11th blast meeting.Mol Microbiol. 2011 Jul;81(1):8-22. doi: 10.1111/j.1365-2958.2011.07694.x. Epub 2011 May 23. Mol Microbiol. 2011. PMID: 21564335 Free PMC article.
References
-
- Padan E, Zilberstein D, Rottenberg H. The proton electrochemical gradient in Escherichia coli cells. Eur J Biochem. 1976;63:533–541. - PubMed
-
- Soksawatmaekhin W, Kuraishi A, Sakata K, Kashiwagi K, Igarashi K. Excretion and uptake of cadaverine by CadB and its physiological functions in Escherichia coli. Mol Microbiol. 2004;51:1401–1412. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
