

Sensorimotor integration in speech processing: computational basis and neural organization
- PMID: 21315253
- PMCID: PMC3057382
- DOI: 10.1016/j.neuron.2011.01.019
Sensorimotor integration in speech processing: computational basis and neural organization
Abstract
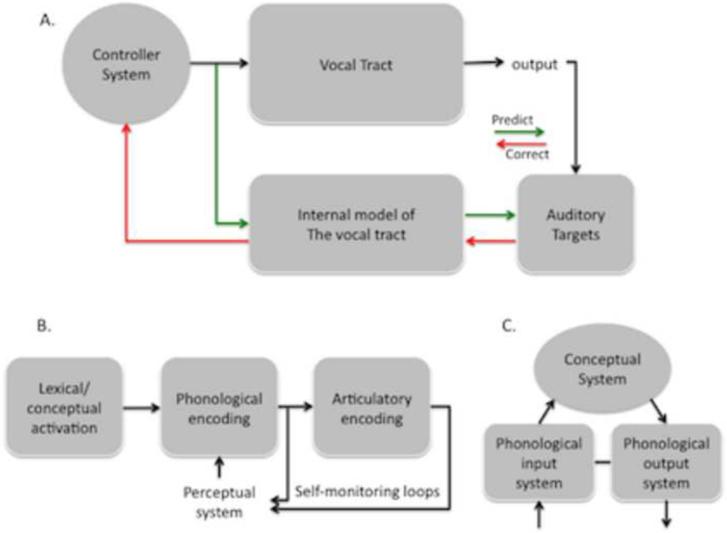
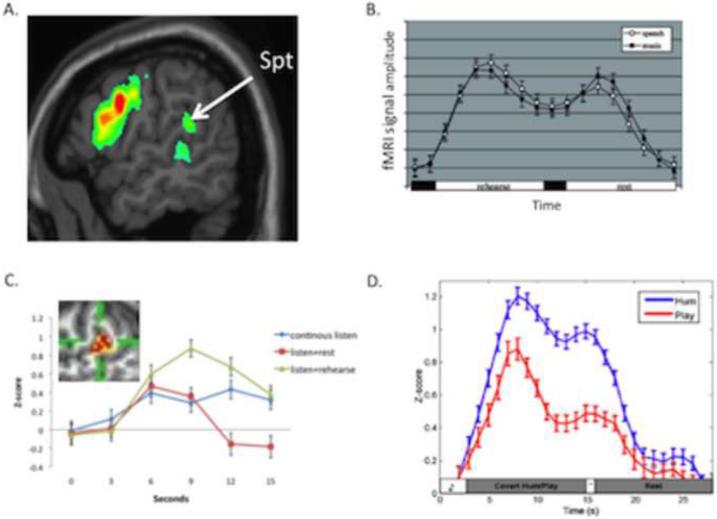
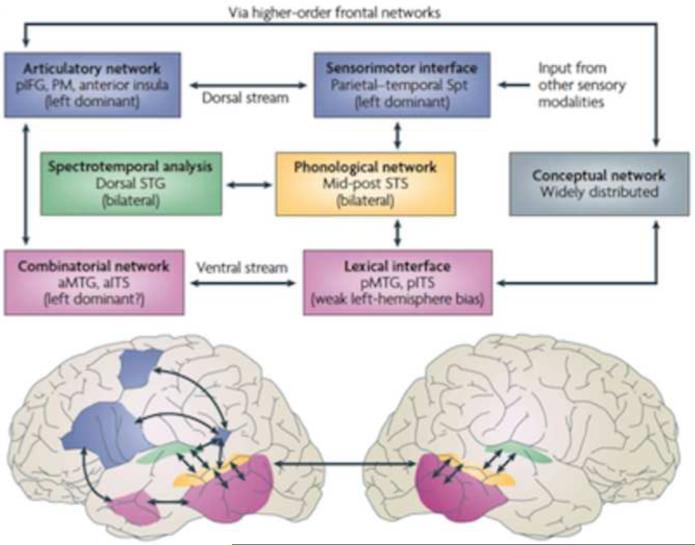
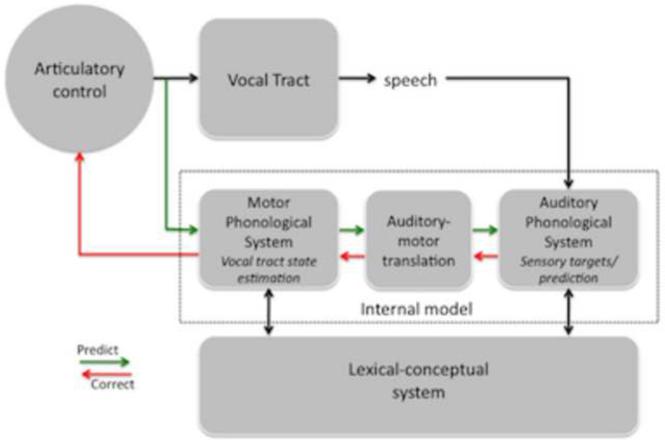
Sensorimotor integration is an active domain of speech research and is characterized by two main ideas, that the auditory system is critically involved in speech production and that the motor system is critically involved in speech perception. Despite the complementarity of these ideas, there is little crosstalk between these literatures. We propose an integrative model of the speech-related "dorsal stream" in which sensorimotor interaction primarily supports speech production, in the form of a state feedback control architecture. A critical component of this control system is forward sensory prediction, which affords a natural mechanism for limited motor influence on perception, as recent perceptual research has suggested. Evidence shows that this influence is modulatory but not necessary for speech perception. The neuroanatomy of the proposed circuit is discussed as well as some probable clinical correlates including conduction aphasia, stuttering, and aspects of schizophrenia.
Copyright © 2011 Elsevier Inc. All rights reserved.
Figures
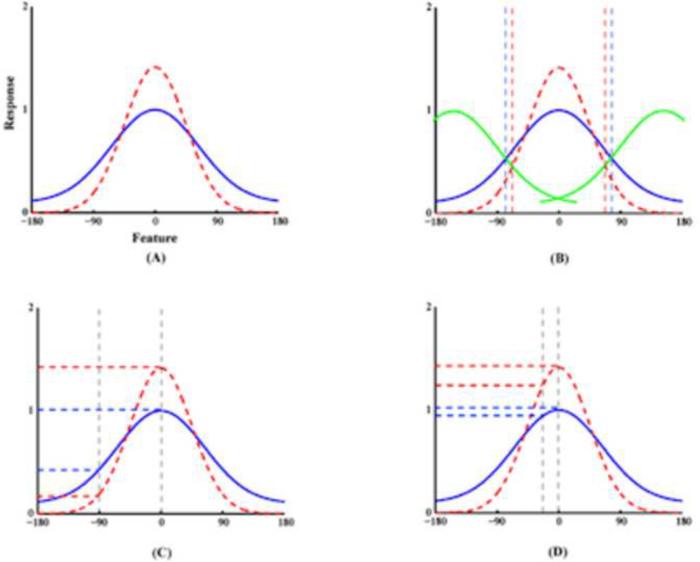
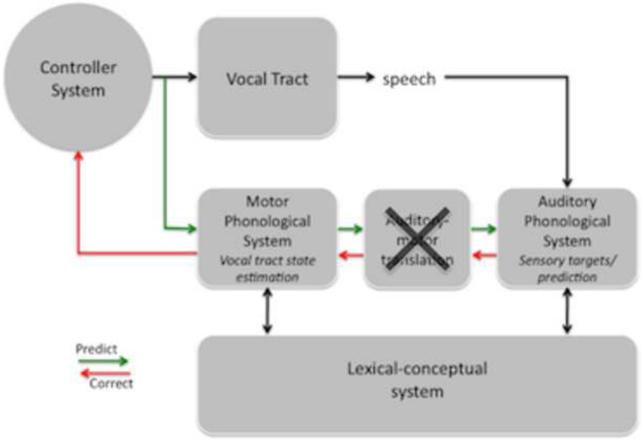
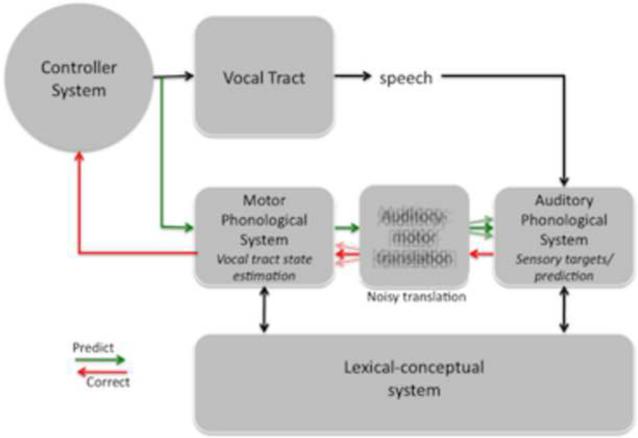
Comment in
-
What is optimal about motor control?Neuron. 2011 Nov 3;72(3):488-98. doi: 10.1016/j.neuron.2011.10.018. Neuron. 2011. PMID: 22078508
References
-
- Anderson JM, Gilmore R, Roper S, Crosson B, Bauer RM, Nadeau S, Beversdorf DQ, Cibula J, Rogish M, III, Kortencamp S, et al. Conduction aphasia and the arcuate fasciculus: A reexamination of the Wernicke-Geschwind model. Brain and Language. 1999;70:1–12. - PubMed
-
- Baldo JV, Klostermann EC, Dronkers NF. It's either a cook or a baker: patients with conduction aphasia get the gist but lose the trace. Brain Lang. 2008;105:134–140. - PubMed
-
- Benson DF, Sheremata WA, Bouchard R, Segarra JM, Price D, Geschwind N. Conduction aphasia: A clincopathological study. Archives of Neurology. 1973;28:339–346. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
