

EZH1 and EZH2 cogovern histone H3K27 trimethylation and are essential for hair follicle homeostasis and wound repair
- PMID: 21317239
- PMCID: PMC3049289
- DOI: 10.1101/gad.2019811
EZH1 and EZH2 cogovern histone H3K27 trimethylation and are essential for hair follicle homeostasis and wound repair
Abstract
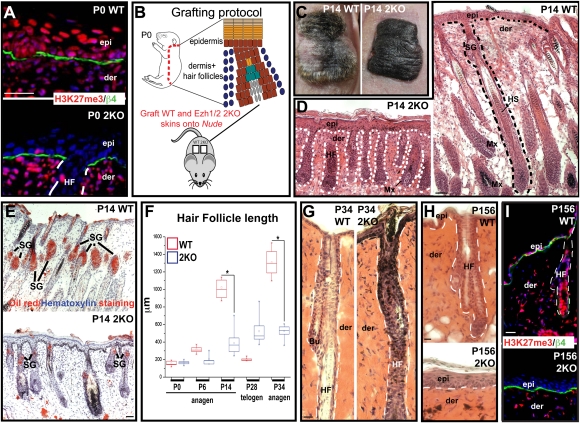
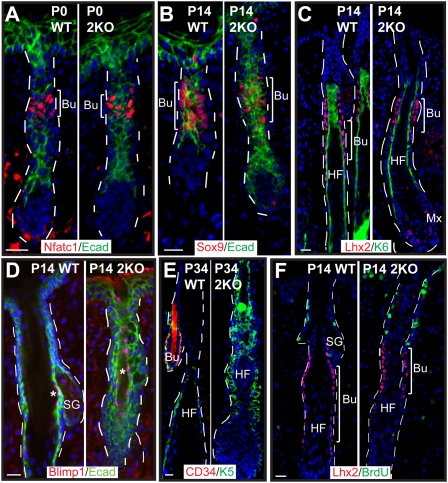
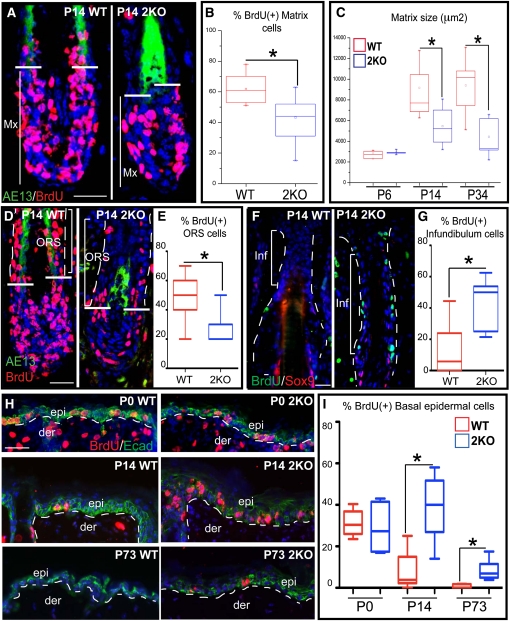
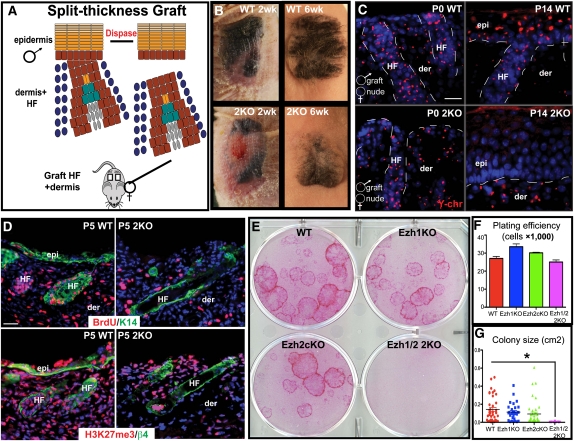
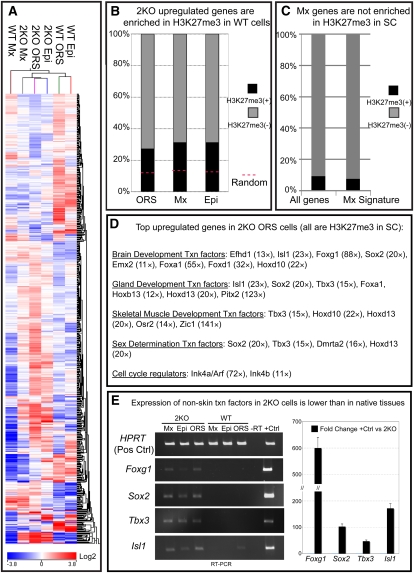
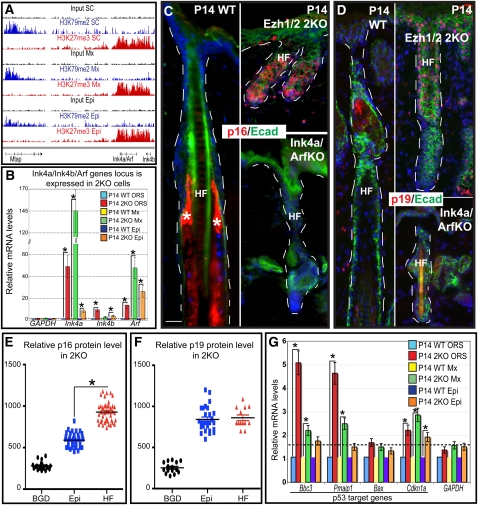
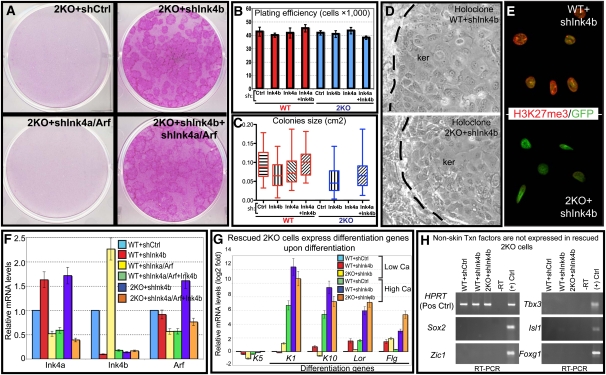
Polycomb protein group (PcG)-dependent trimethylation on H3K27 (H3K27me3) regulates identity of embryonic stem cells (ESCs). How H3K27me3 governs adult SCs and tissue development is unclear. Here, we conditionally target H3K27 methyltransferases Ezh2 and Ezh1 to address their roles in mouse skin homeostasis. Postnatal phenotypes appear only in doubly targeted skin, where H3K27me3 is abolished, revealing functional redundancy in EZH1/2 proteins. Surprisingly, while Ezh1/2-null hair follicles (HFs) arrest morphogenesis and degenerate due to defective proliferation and increased apoptosis, epidermis hyperproliferates and survives engraftment. mRNA microarray studies reveal that, despite these striking phenotypic differences, similar genes are up-regulated in HF and epidermal Ezh1/2-null progenitors. Featured prominently are (1) PcG-controlled nonskin lineage genes, whose expression is still significantly lower than in native tissues, and (2) the PcG-regulated Ink4a/Inkb/Arf locus. Interestingly, when EZH1/2 are absent, even though Ink4a/Arf/Ink4b genes are fully activated in HF cells, they are only partially so in epidermal progenitors. Importantly, transduction of Ink4b/Ink4a/Arf shRNAs restores proliferation/survival of Ezh1/2-null HF progenitors in vitro, pointing toward the relevance of this locus to the observed HF phenotypes. Our findings reveal new insights into Polycomb-dependent tissue control, and provide a new twist to how different progenitors within one tissue respond to loss of H3K27me3.
Figures
References
-
- Barski A, Cuddapah S, Cui K, Roh TY, Schones DE, Wang Z, Wei G, Chepelev I, Zhao K 2007. High-resolution profiling of histone methylations in the human genome. Cell 129: 823–837 - PubMed
-
- Blanpain C, Lowry WE, Geoghegan A, Polak L, Fuchs E 2004. Self-renewal, multipotency, and the existence of two cell populations within an epithelial stem cell niche. Cell 118: 635–648 - PubMed
-
- Boyer LA, Plath K, Zeitlinger J, Brambrink T, Medeiros LA, Lee TI, Levine SS, Wernig M, Tajonar A, Ray MK, et al. 2006. Polycomb complexes repress developmental regulators in murine embryonic stem cells. Nature 441: 349–353 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous