

Mitosis in the human malaria parasite Plasmodium falciparum
- PMID: 21317311
- PMCID: PMC3127633
- DOI: 10.1128/EC.00314-10
Mitosis in the human malaria parasite Plasmodium falciparum
Abstract
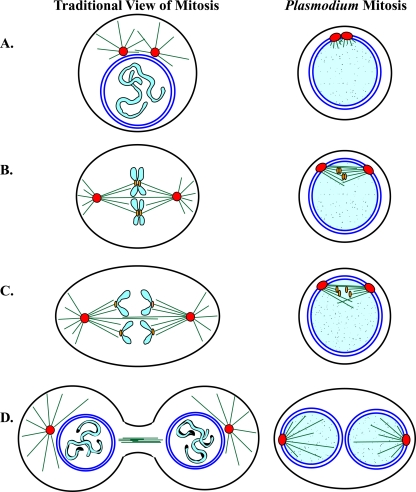
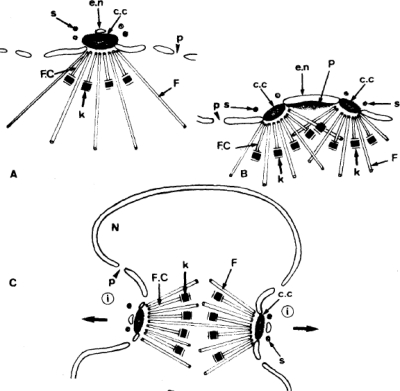
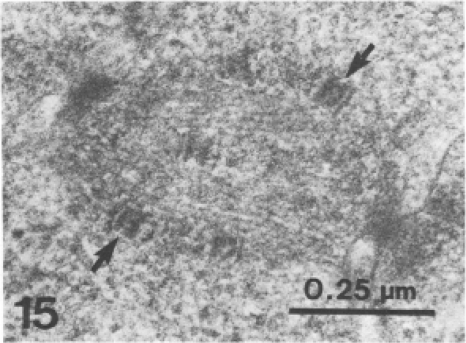
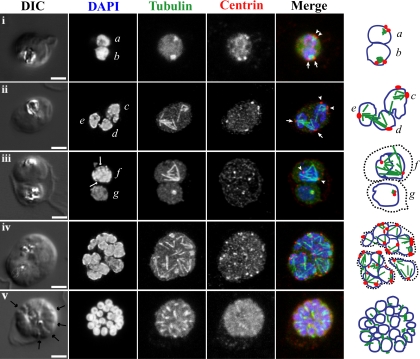
Malaria is caused by intraerythrocytic protozoan parasites belonging to Plasmodium spp. (phylum Apicomplexa) that produce significant morbidity and mortality, mostly in developing countries. Plasmodium parasites have a complex life cycle that includes multiple stages in anopheline mosquito vectors and vertebrate hosts. During the life cycle, the parasites undergo several cycles of extreme population growth within a brief span, and this is critical for their continued transmission and a contributing factor for their pathogenesis in the host. As with other eukaryotes, successful mitosis is an essential requirement for Plasmodium reproduction; however, some aspects of Plasmodium mitosis are quite distinct and not fully understood. In this review, we will discuss the current understanding of the architecture and key events of mitosis in Plasmodium falciparum and related parasites and compare them with the traditional mitotic events described for other eukaryotes.
Figures
References
-
- Adisa A., et al. 2007. Re-assessing the locations of components of the classical vesicle-mediated trafficking machinery in transfected Plasmodium falciparum. Int. J. Parasitol. 37:1127–1141 - PubMed
-
- Aikawa M. 1971. Parasitological review. Plasmodium: the fine structure of malarial parasites. Exp. Parasitol. 30:284–320 - PubMed
-
- Alano P., Billker O. 2005. Gametocytes and gametes, p. 191–219 In Sherman I. W. (ed.), Molecular approaches to malaria. ASM Press, Washington, DC
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
