

Evolution and functional diversification of MIRNA genes
- PMID: 21317375
- PMCID: PMC3077775
- DOI: 10.1105/tpc.110.082784
Evolution and functional diversification of MIRNA genes
Abstract
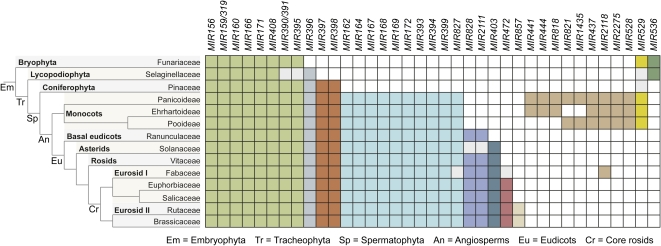
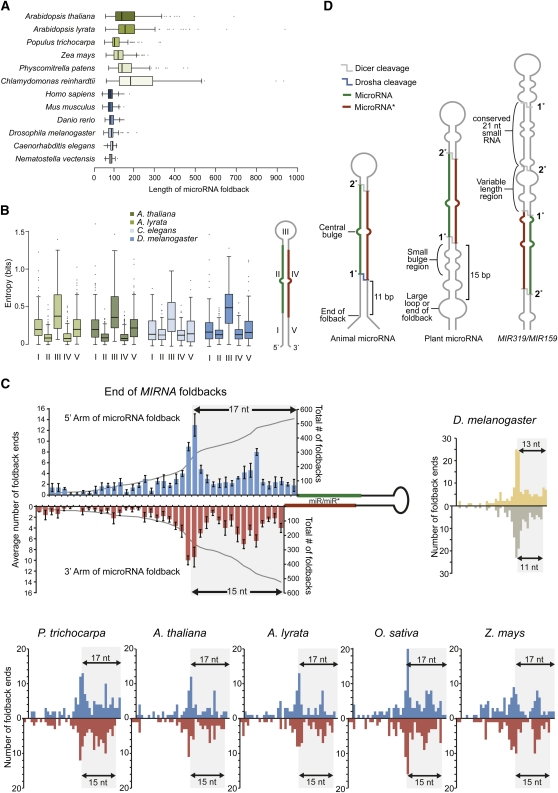
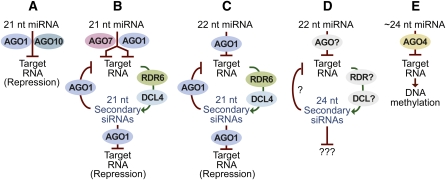
MicroRNAs (miRNAs) are small regulatory RNAs found in diverse eukaryotic lineages. In plants, a minority of annotated MIRNA gene families are conserved between plant families, while the majority are family- or species-specific, suggesting that most known MIRNA genes arose relatively recently in evolutionary time. Given the high proportion of young MIRNA genes in plant species, new MIRNA families are likely spawned and then lost frequently. Unlike highly conserved, ancient miRNAs, young miRNAs are often weakly expressed, processed imprecisely, lack targets, and display patterns of neutral variation, suggesting that young MIRNA loci tend to evolve neutrally. Genome-wide analyses from several plant species have revealed that variation in miRNA foldback expression, structure, processing efficiency, and miRNA size have resulted in the unique functionality of MIRNA loci and resulting miRNAs. Additionally, some miRNAs have evolved specific properties and functions that regulate other transcriptional or posttranscriptional silencing pathways. The evolution of miRNA processing and functional diversity underscores the dynamic nature of miRNA-based regulation in complex regulatory networks.
Figures
References
-
- Allen E., Howell M.D. (2010). miRNAs in the biogenesis of trans-acting siRNAs in higher plants. Semin. Cell Dev. Biol. 21: 798–804 - PubMed
-
- Allen E., Xie Z., Gustafson A.M., Carrington J.C. (2005). microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121: 207–221 - PubMed
-
- Allen E., Xie Z., Gustafson A.M., Sung G.H., Spatafora J.W., Carrington J.C. (2004). Evolution of microRNA genes by inverted duplication of target gene sequences in Arabidopsis thaliana. Nat. Genet. 36: 1282–1290 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
