

Stimulation of olfactory ensheathing cell motility enhances olfactory axon growth
- PMID: 21318262
- PMCID: PMC11115065
- DOI: 10.1007/s00018-011-0630-9
Stimulation of olfactory ensheathing cell motility enhances olfactory axon growth
Abstract
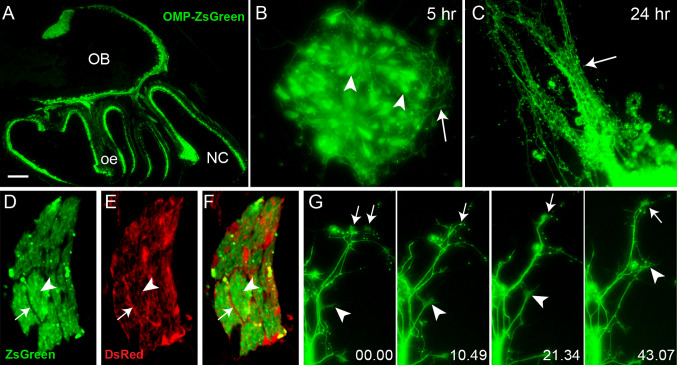
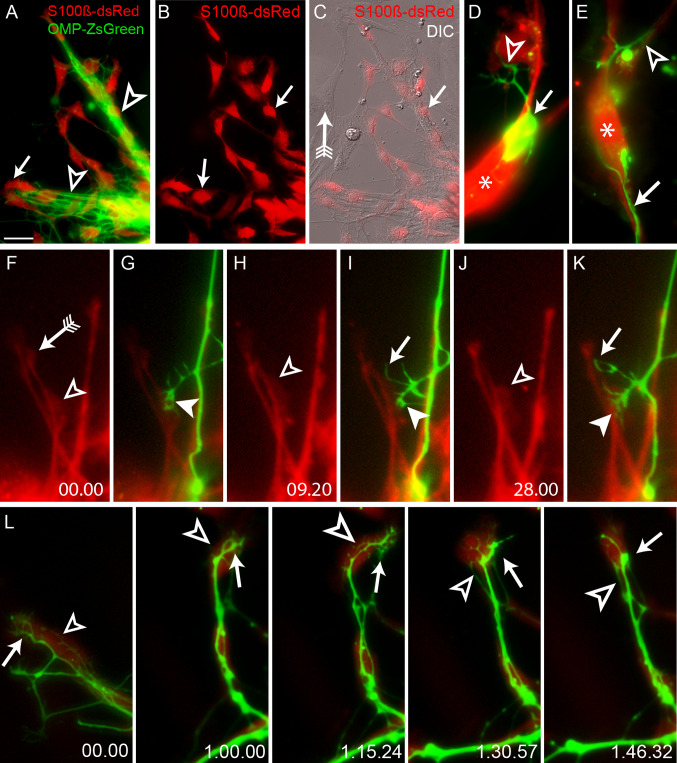
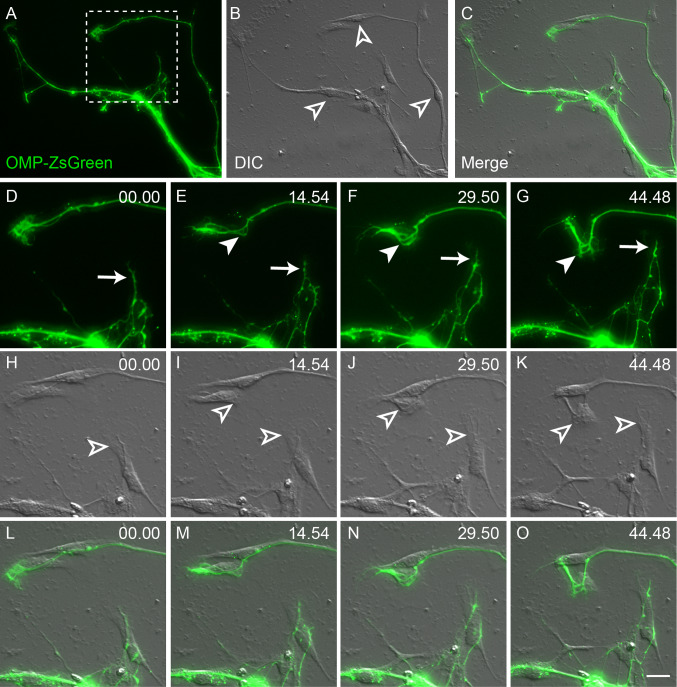
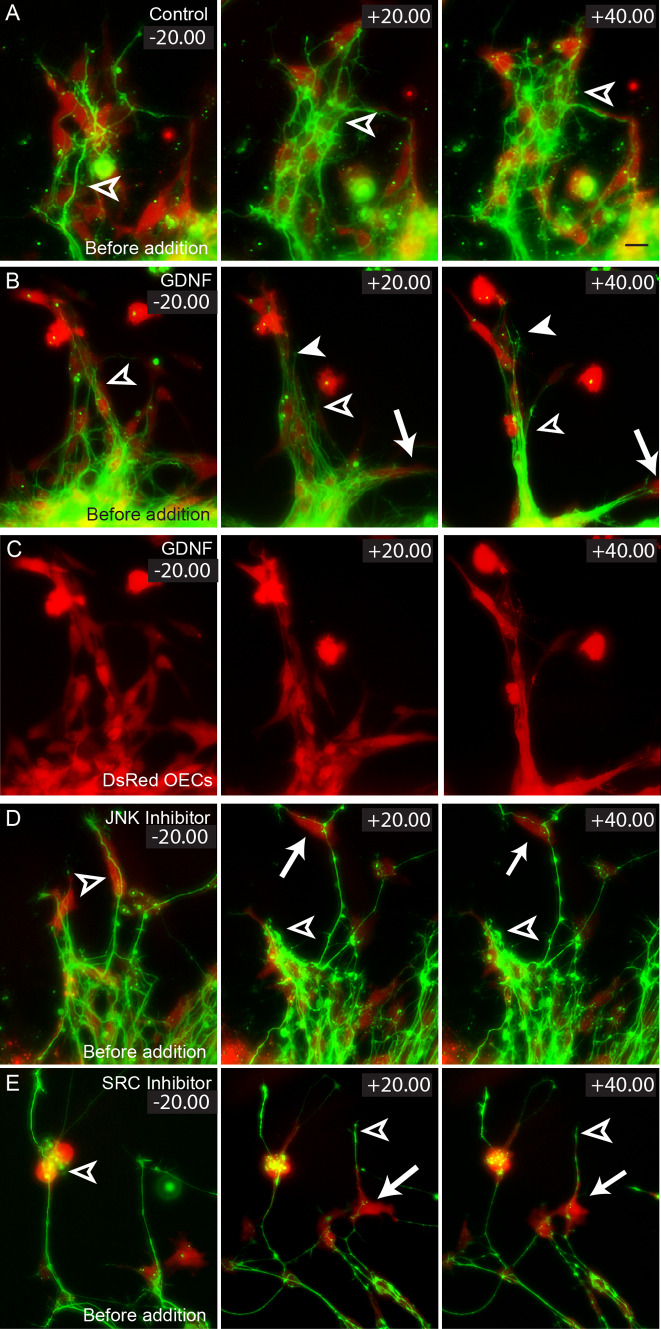
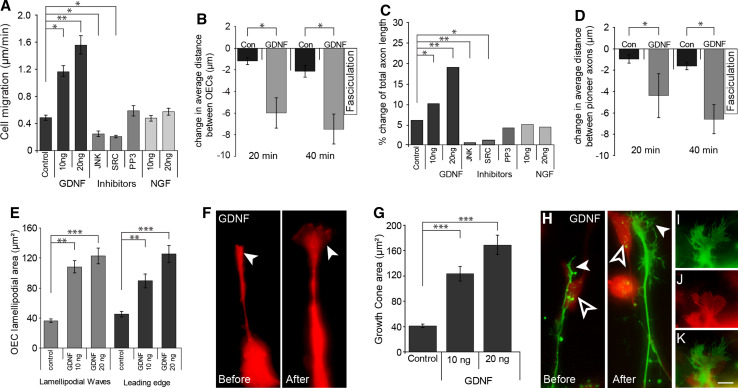
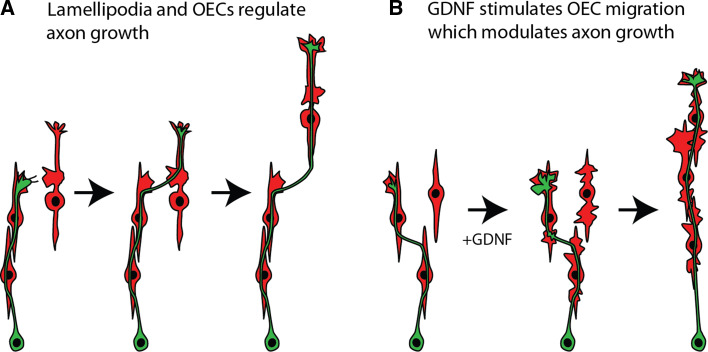
Axons of primary olfactory neurons are intimately associated with olfactory ensheathing cells (OECs) from the olfactory epithelium until the final targeting of axons within the olfactory bulb. However, little is understood about the nature and role of interactions between OECs and axons during development of the olfactory nerve pathway. We have used high resolution time-lapse microscopy to examine the growth and interactions of olfactory axons and OECs in vitro. Transgenic mice expressing fluorescent reporters in primary olfactory axons (OMP-ZsGreen) and ensheathing cells (S100ß-DsRed) enabled us to selectively analyse these cell types in explants of olfactory epithelium. We reveal here that rather than providing only a permissive substrate for axon growth, OECs play an active role in modulating the growth of pioneer olfactory axons. We show that the interactions between OECs and axons were dependent on lamellipodial waves on the shaft of OEC processes. The motility of OECs was mediated by GDNF, which stimulated cell migration and increased the apparent motility of the axons, whereas loss of OECs via laser ablation of the cells inhibited olfactory axon outgrowth. These results demonstrate that the migration of OECs strongly regulates the motility of axons and that stimulation of OEC motility enhances axon extension and growth cone activity.
© Springer Basel AG 2011
Figures

References
-
- Chung RS, Woodhouse A, Fung S, Dickson TC, West AK, Vickers JC, Chuah MI. Olfactory ensheathing cells promote neurite sprouting of injured axons in vitro by direct cellular contact and secretion of soluble factors. Cell Mol Life Sci. 2004;61:1238–1245. doi: 10.1007/s00018-004-4026-y. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
