

Sequential stages in the age-dependent gradual formation and accumulation of tubular aggregates in fast twitch muscle fibers: SERCA and calsequestrin involvement
- PMID: 21318331
- PMCID: PMC3260353
- DOI: 10.1007/s11357-011-9211-y
Sequential stages in the age-dependent gradual formation and accumulation of tubular aggregates in fast twitch muscle fibers: SERCA and calsequestrin involvement
Abstract
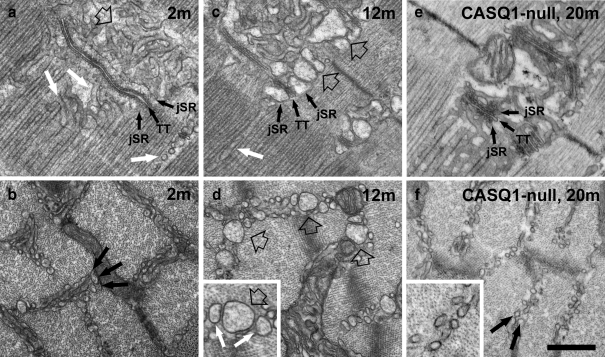
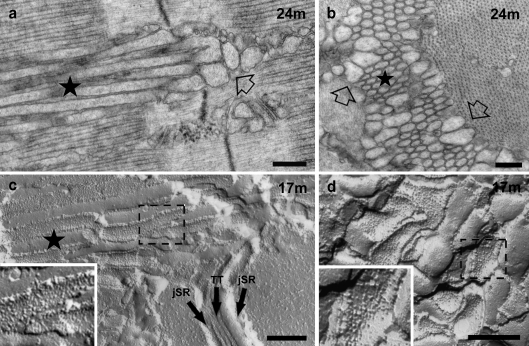
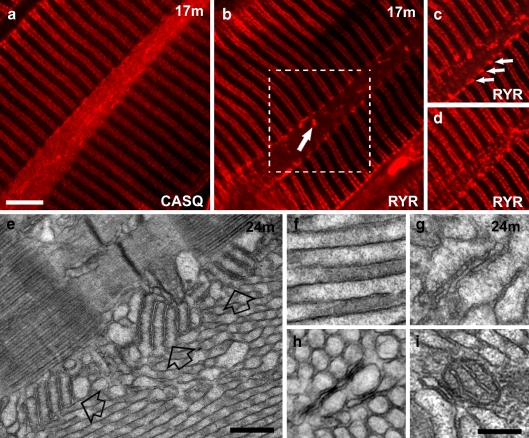
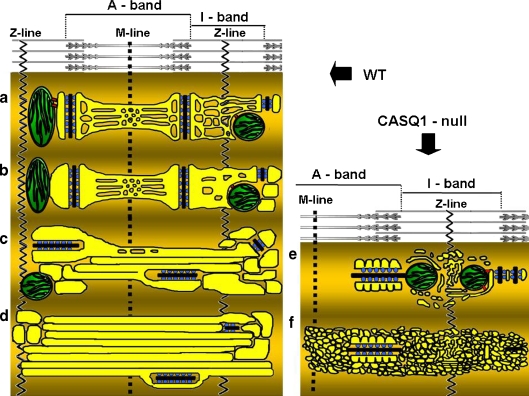
Tubular aggregates (TAs), ordered arrays of elongated sarcoplasmic reticulum (SR) tubules, are present in skeletal muscle from patients with myopathies and are also experimentally induced by extreme anoxia. In wild-type mice TAs develop in a clear age-, sex- (male), and fiber type- (fast twitch) dependence. However, the events preceding the appearance of TAs have not been explored. We investigated the sequential stages leading to the initial appearance and maturation of TAs in EDL from male mice. TAs' formation requires two temporally distinct steps that operate via different mechanisms. Initially (before 1 year of age), the SR Ca(2+) binding protein calsequestrin (CASQ) accumulates specifically at the I band level causing swelling of free SR cisternae. In the second stage, the enlarged SR sacs at the I band level extend into multiple, longitudinally oriented tubules with a full complement of sarco(endo)plasmic reticulum Ca(2+) ATPases (SERCA) in the membrane and CASQ in the lumen. Tubules gradually acquire a regular cylindrical shape and uniform size apparently in concert with partial crystallization of SERCA. Multiple, small TAs associate to form fewer mature TAs of very large size. Interestingly, in fibers from CASQ1-knockout mice abnormal aggregates of SR tubules have different conformation and never develop into ordered aggregates of straight cylinders, possibly due to lack of CASQ accumulation. We conclude that TAs do not arise abruptly but are the final result of a gradually changing SR architecture and we suggest that the crystalline ATPase within the aggregates may be inactive.
Figures



References
-
- Agbulut O, Destombes J, Thiesson D, Butler-Browne G. Age-related appearance of tubular aggregates in the skeletal muscle of almost all male inbred mice. Histochem Cell Biol. 2000;114(6):477–481. - PubMed
-
- Airey JA, Beck CF, Murakami K, Tanksley SJ, Deerinck TJ, Ellisman MH, Sutko JL (1990) Identification and localization of two triad junctional foot protein isoforms in mature avian fast-twitch skeletal muscle. J Biol Chem 265:14187–14194 - PubMed
-
- Augusto V, Padovani CR, Campos GER. Skeletal muscle fiber types in c57bl6j mice. Braz J morphol Sci. 2004;21(2):89–94.
-
- Boncompagni S, d’Amelio L, Fulle S, Fano G, Protasi F. Progressive disorganization of the excitation-contraction coupling apparatus in aging human skeletal muscle as revealed by electron microscopy: A possible role in the decline of muscle performance. J Gerontol A Biol Sci Med Sci. 2006;61(10):995–1008. doi: 10.1093/gerona/61.10.995. - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous