

Review
doi: 10.1089/hum.2010.204.
Epub 2011 Apr 6.
Adeno-associated viral vectors for mapping, monitoring, and manipulating neural circuits
Affiliations
- PMID: 21319997
- PMCID: PMC3107581
- DOI: 10.1089/hum.2010.204
Item in Clipboard
Review
Adeno-associated viral vectors for mapping, monitoring, and manipulating neural circuits
Hum Gene Ther.
2011 Jun.
Abstract
Understanding the structure and function of neural circuits is central is neuroscience research. To address the associated questions, new genetically encoded tools have been developed for mapping, monitoring, and manipulating neurons. Essential to implementation of these tools is their selective delivery to defined neuronal populations in the brain. This has been facilitated by recent improvements in cell type-specific transgene expression using recombinant adeno-associated viral vectors. Here, we highlight these developments and discuss areas for improvement that could further expand capabilities for neural circuit analysis.
Figures
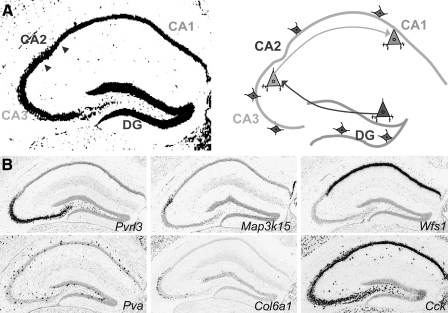
Marker genes specify circuit nodes in hippocampal circuitry. (A) Connections between anatomical regions of the hippocampus. In hippocampal circuits, granule cells located in the dentate gyrus (DG) send axonal projections to the CA3 region. The pyramidal cells in CA3 send axonal projections to CA1 neurons. Interspersed throughout the hippocampus are multiple classes of local inhibitory interneurons (diamond shaped in scheme). (B) Gene expression patterns label neurons selectively in the three main anatomical domains of the hippocampus. Pvrl3 marks CA3; Col6a1 marks a subset of CA3 neurons; Map3k15 marks CA2; Wfs1 marks CA1. Two inhibitory interneuron populations interspersed throughout the hippocampus can be distinguished by the expression of Pva or Cck. Within the hippocampal region, these genes mark neurons in distinct anatomical domains, however the expressed genes are not exclusive to these hippocampal regions, as they are also expressed in other anatomical domains of the brain. Images taken from: Allen Mouse Brain Atlas. Seattle (WA): Allen Institute for Brain Science. Copyright, 2009. (Lein et al., 2007) Available from: http://mouse.brain-map.org .
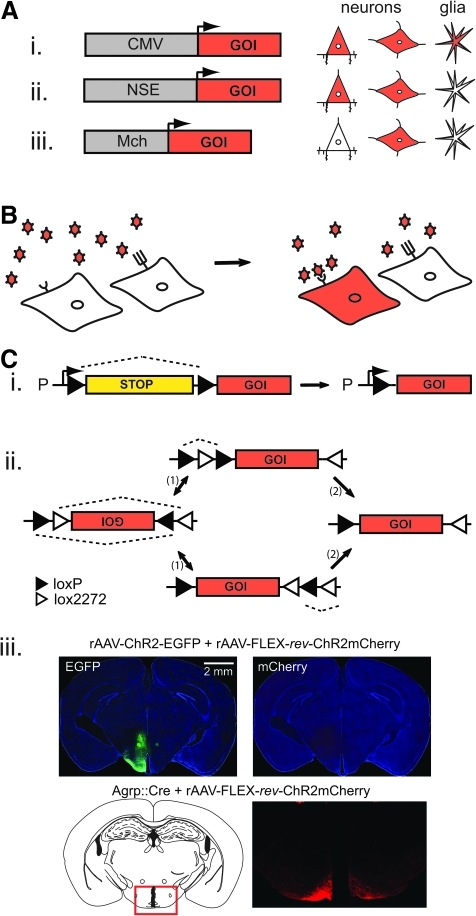
Strategies for cell type–selective transgene expression with recombinant adeno-associated virus (rAAV). (A) Promoter specificity. Schematic showing cell-type specificity of short promoter fragments in rAAV. The cytomegalovirus (CMV) promoter drives expression nonselectively in neurons and glia, while the neuron-specific enolase (NSE) promoter is selective for neurons, and the melanin-concentrating hormone (MCH) promoter targets expression to a subpopulation of neurons in the lateral hypothalamus. (B) Tropism specificity. AAV capsid proteins lead to differential infectivity between cell populations based on recognition between capsids and cell surface receptors. (C) Cre-dependent intersectional genetic switches. (i) Insertion of a 1–2 kb stop cassette after the promoter (P) and upstream of a gene of interest (GOI). The stop cassette is flanked by two loxP sites which are the recognition sequences that recombine in the presence of Cre. After recombination, the stop cassette is excised (one loxP site remains) and transcription of the GOI proceeds. (ii) Use of a flip-excision (FLEX) switch to activate gene expression specifically in the presence of Cre-recombinase. The GOI is inverted between two pairs of heterotypic, antiparallel lox-type recombination sites, which first undergo an inversion of the coding sequence, followed by excision of two sites, leading to one of each orthogonal recombination site oppositely oriented and incapable of further recombination. (iii) Demonstration of Cre-dependent transgene expression using a FLEX switch. The top panels show tissue from a wild-type mouse brain co-injected with both a rAAV2/1-ChR2-EGFP virus and a rAAV2/1-FLEX-rev-ChR2-mCherry virus. Expression of EGFP is broadly observed while mCherry is not expressed in the absence of Cre. Injection of rAAV2/1-FLEX-rev-ChR2-mCherry into an Agrp::Cre mouse (bottom panel) shows expression of the mCherry signal only in the arcuate nucleus, which is the sole location of AGRP neurons in the brain. Adapted from Journal of Neuroscience (Atasoy et al., 2008), copyright 2008.
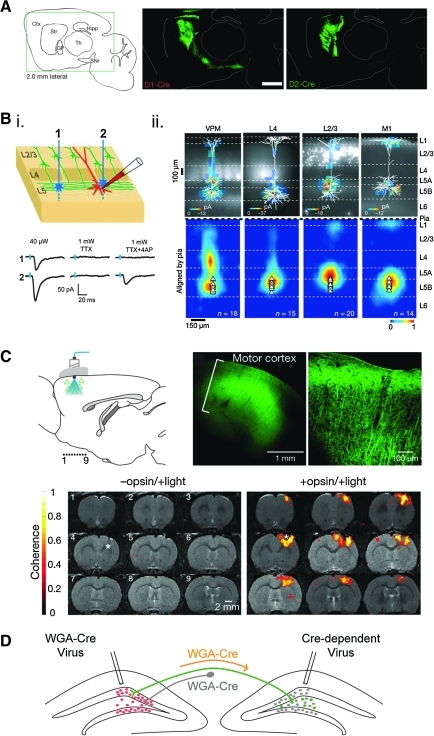
Applications of rAAV for neural circuit analysis. (A) Selective, Cre-dependent viral-mediated expression of YFP in two distinct populations of striatal neurons. D1R-expressing neurons project to the substantia nigra pars reticulata (SNr), D2R-expressing neurons project to the globus pallidus (GP). These experiments highlight discrete projection patterns for molecularly defined neurons occupying the same anatomical, indicating distinct neuronal functions. Reprinted by permission from Macmillan Publishers Ltd: Nature (Kravitz et al., 2010), copyright 2010. www.nature.com . (B) Mapping the subcellular distribution of inputs onto pyramidal neurons in the cortex using sCRACM. (i) The top panel shows two photostimulation configurations, where the postsynaptic neuron (red) is being recorded from and the blue stars indicate spots where the ChR2 expressing axons (green) are stimulated. The bottom panel provides representative traces of excitatory postsynaptic currents evoked by photostimulation corresponding to the locations indicated in the sketch. In the absence of the voltage-gated sodium channel blocker tetrodotoxin (TTX), both stimulation sites evoke synaptic responses, but these are both blocked with TTX, which prevents action potentials. Addition of the potassium channel blocker 4-aminopyridine (4-AP) renders ChR2 activation at a synaptic connection sufficient to drive release and evoke a synaptic response. Blue tickmarks indicate the laser pulse. Laser power is also shown for these conditions. (ii) Shown in the top panel are examples of sCRACM maps overlaid on somato-dendritic reconstructions. The fluorescent neurons are ChR2-positive neurons that are being mapped onto a pyramidal neuron. The bottom panel shows group averages of the inputs onto the post-synaptic neuron. Reprinted by permission from Macmillan Publishers Ltd: Nature (Petreanu et al., 2009), copyright 2009. www.nature.com . (C) Functional magnetic resonance imaging (fMRI) with blood oxygenation level detection (BOLD) signal detects activation of neurons resulting from cortical neuron stimulation. Diagram shows cortical neurons transduced by a virus expressing the ChR2-EYFP transgene as triangles and the path of the photostimulation in blue. Adjacent images are of ChR2-EYFP expressing neurons in the motor cortex. In brain slices that lack the ChR2 opsin, negligible levels of BOLD activation are observed; however, when AAV5-ChR2 is expressed in the cortex (+opsin), BOLD activation is observed. Reprinted by permission from Macmillan Publishers Ltd: Nature (Lee et al., 2010), copyright 2010. www.nature.com (D) Target region specificity. Injection of a Cre-dependent virus to infect neurons in one region of the central nervous system can be combined with an injection of a virus expressing wheat germ agglutinin (WGA)-Cre in a known anatomical target region of a subset of neurons in the original infection region. The fusion of WGA to the Cre recombinase promotes retrograde transport from the axon terminals of all neurons contributing to that projection field back to the cell body. Once at the nucleus, the Cre-recombinase unmasks the expression of the gene of interest (green fluorescent protein [GFP] in this case) of only the neurons in the original infection field that project to the appropriate target region. Reprinted by permission from Elsevier: Cell (Gradinaru et al., 2010), copyright 2010. www.sciencedirect.com/science/journal/00928674
Similar articles
-
Strategies for targeting primate neural circuits with viral vectors.J Neurophysiol. 2016 Jul 1;116(1):122-34. doi: 10.1152/jn.00087.2016. Epub 2016 Apr 6. J Neurophysiol. 2016. PMID: 27052579 Free PMC article. Review.
-
Local and retrograde gene transfer into primate neuronal pathways via adeno-associated virus serotype 8 and 9.Neuroscience. 2011 Oct 13;193:249-58. doi: 10.1016/j.neuroscience.2011.06.080. Epub 2011 Jul 18. Neuroscience. 2011. PMID: 21782903
-
Translational PET applications for brain circuit mapping with transgenic neuromodulation tools.Pharmacol Biochem Behav. 2021 May;204:173147. doi: 10.1016/j.pbb.2021.173147. Epub 2021 Feb 4. Pharmacol Biochem Behav. 2021. PMID: 33549570 Free PMC article. Review.
-
Manipulating gene expression in projection-specific neuronal populations using combinatorial viral approaches.Curr Protoc Neurosci. 2013;65(435):4.35.1-20. doi: 10.1002/0471142301.ns0435s65. Curr Protoc Neurosci. 2013. PMID: 25429312 Free PMC article.
-
Use of Adeno-Associated and Herpes Simplex Viral Vectors for In Vivo Neuronal Expression in Mice.Curr Protoc Neurosci. 2015 Oct 1;73:4.37.1-4.37.31. doi: 10.1002/0471142301.ns0437s73. Curr Protoc Neurosci. 2015. PMID: 26426386 Free PMC article.
Cited by
-
Programmable Assembly of Adeno-Associated Virus-Antibody Composites for Receptor-Mediated Gene Delivery.Bioconjug Chem. 2020 Apr 15;31(4):1093-1106. doi: 10.1021/acs.bioconjchem.9b00790. Epub 2019 Dec 20. Bioconjug Chem. 2020. PMID: 31809024 Free PMC article.
-
Off-Target Expression of Cre-Dependent Adeno-Associated Viruses in Wild-Type C57BL/6J Mice.eNeuro. 2021 Nov 24;8(6):ENEURO.0363-21.2021. doi: 10.1523/ENEURO.0363-21.2021. Print 2021 Nov-Dec. eNeuro. 2021. PMID: 34785571 Free PMC article.
-
Cre-dependent selection yields AAV variants for widespread gene transfer to the adult brain.Nat Biotechnol. 2016 Feb;34(2):204-9. doi: 10.1038/nbt.3440. Epub 2016 Feb 1. Nat Biotechnol. 2016. PMID: 26829320 Free PMC article.
-
Neuromodulation in Chemosensory Pathways.Chem Senses. 2017 Jun 1;42(5):375-379. doi: 10.1093/chemse/bjx014. Chem Senses. 2017. PMID: 28379355 Free PMC article. Review.
-
CREATEd viruses go global.Nat Neurosci. 2017 Jul 26;20(8):1041-1042. doi: 10.1038/nn.4600. Nat Neurosci. 2017. PMID: 28745726 No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources