

Molecular interaction and functional regulation of connexin50 gap junctions by calmodulin
- PMID: 21320072
- PMCID: PMC3351833
- DOI: 10.1042/BJ20101726
Molecular interaction and functional regulation of connexin50 gap junctions by calmodulin
Abstract
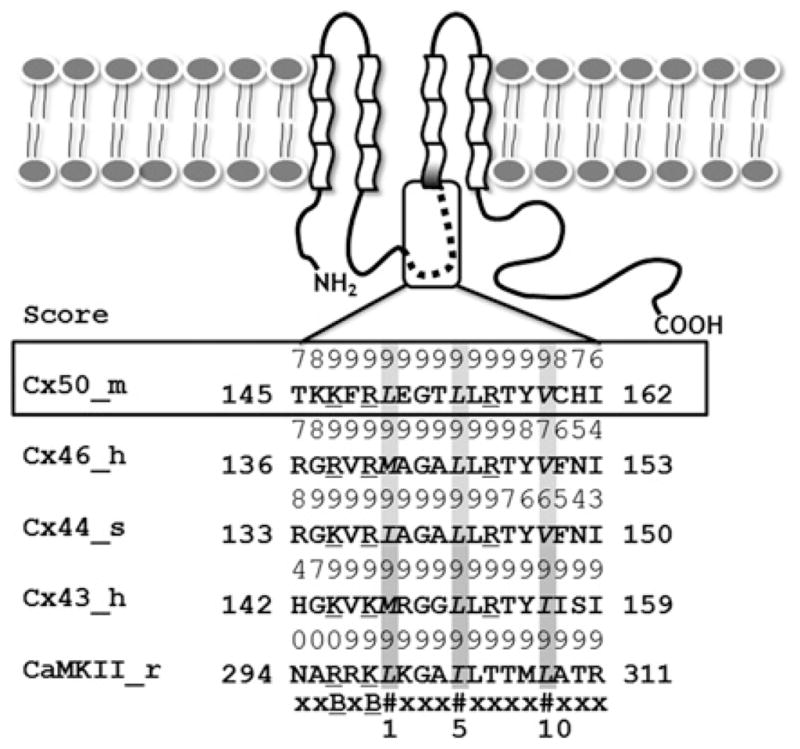
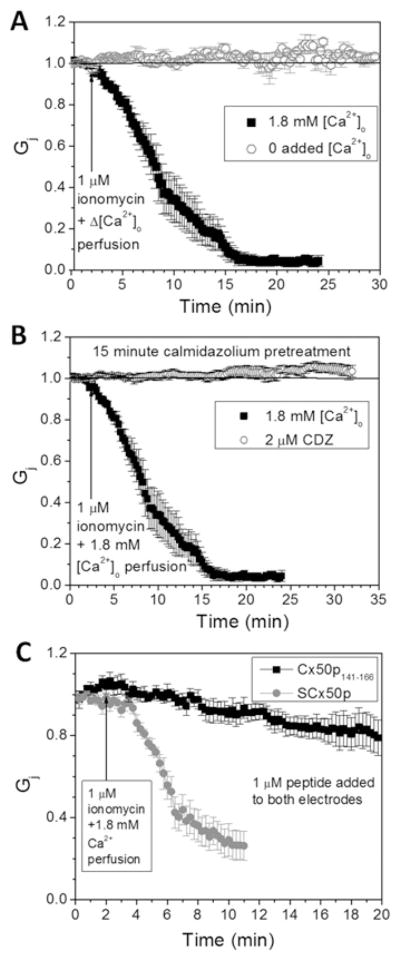
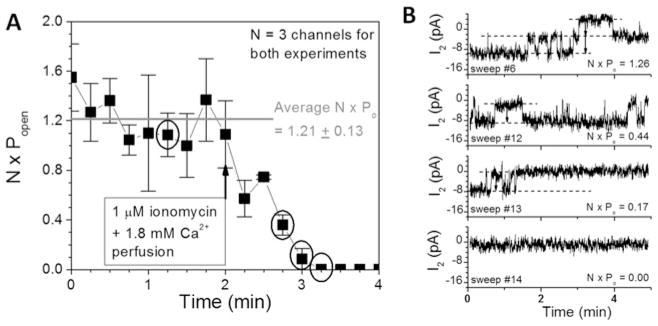
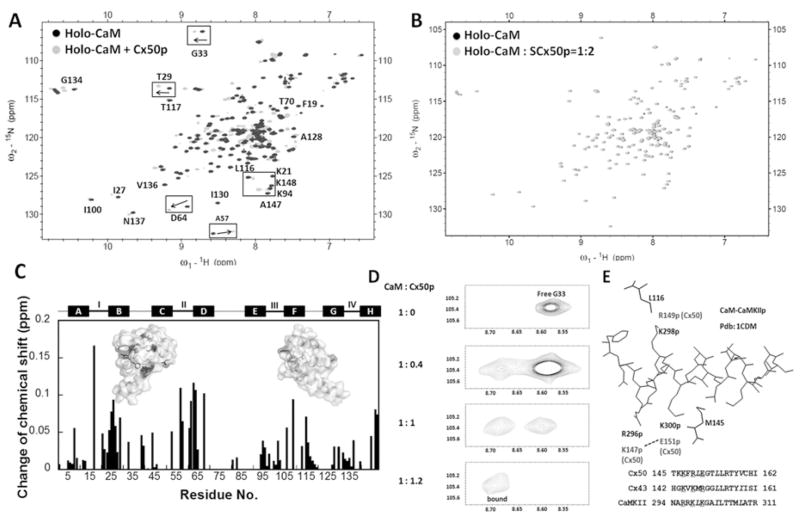
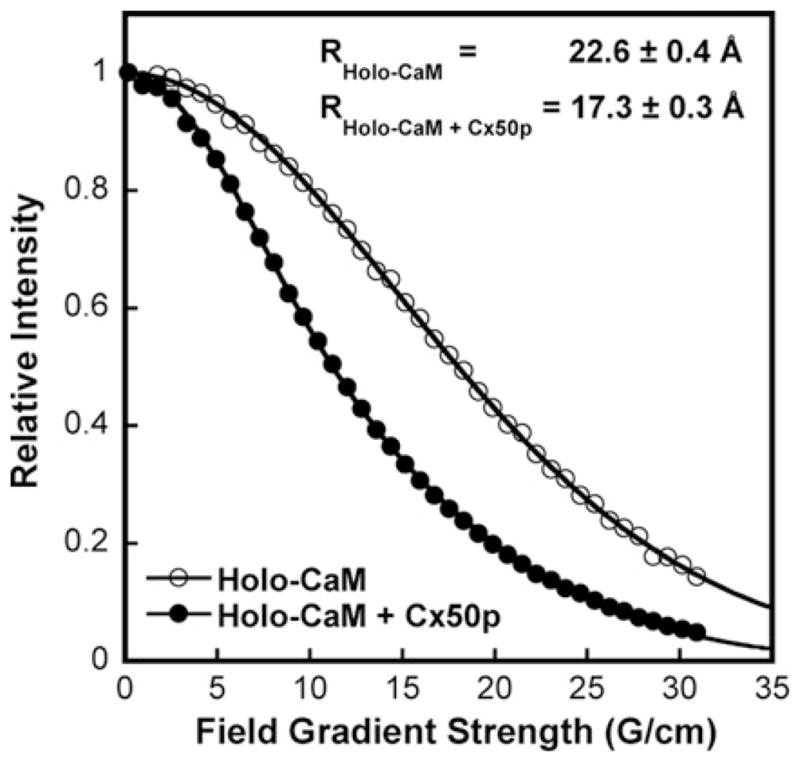
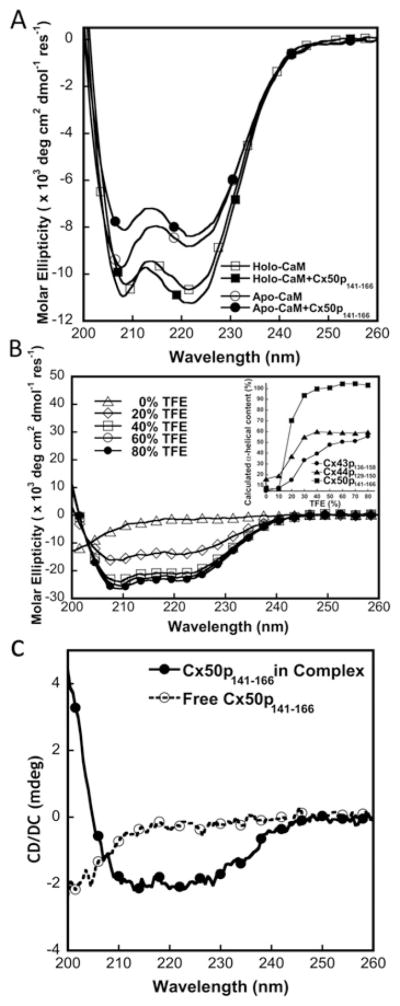
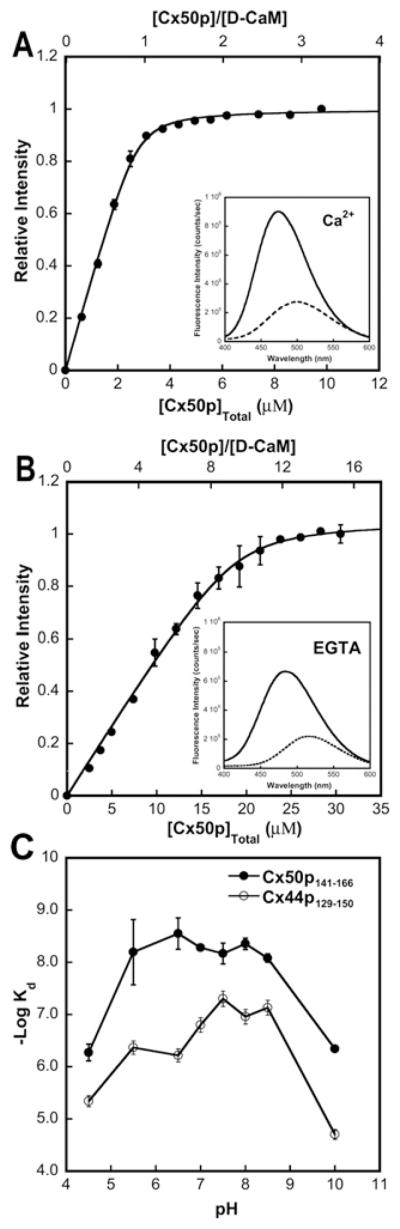
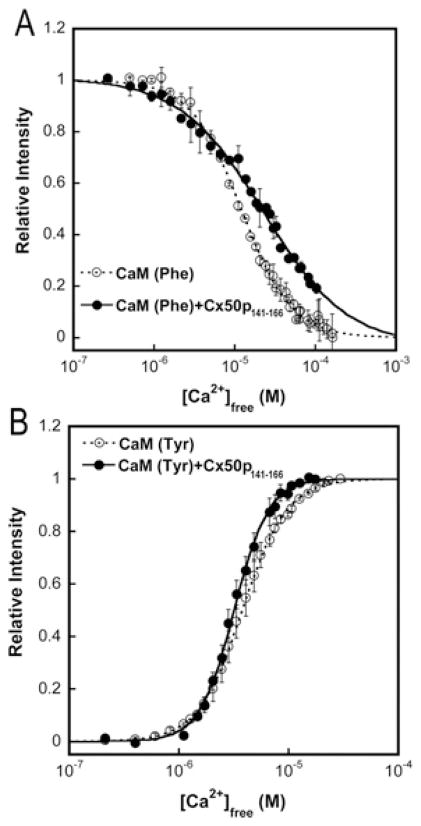
Cx50 (connexin50), a member of the α-family of gap junction proteins expressed in the lens of the eye, has been shown to be essential for normal lens development. In the present study, we identified a CaMBD [CaM (calmodulin)-binding domain] (residues 141-166) in the intracellular loop of Cx50. Elevations in intracellular Ca2+ concentration effected a 95% decline in gj (junctional conductance) of Cx50 in N2a cells that is likely to be mediated by CaM, because inclusion of the CaM inhibitor calmidazolium prevented this Ca2+-dependent decrease in gj. The direct involvement of the Cx50 CaMBD in this Ca2+/CaM-dependent regulation was demonstrated further by the inclusion of a synthetic peptide encompassing the CaMBD in both whole-cell patch pipettes, which effectively prevented the intracellular Ca2+-dependent decline in gj. Biophysical studies using NMR and fluorescence spectroscopy reveal further that the peptide stoichiometrically binds to Ca2+/CaM with an affinity of ~5 nM. The binding of the peptide expanded the Ca2+-sensing range of CaM by increasing the Ca2+ affinity of the C-lobe of CaM, while decreasing the Ca2+ affinity of the N-lobe of CaM. Overall, these results demonstrate that the binding of Ca2+/CaM to the intracellular loop of Cx50 is critical for mediating the Ca2+-dependent inhibition of Cx50 gap junctions in the lens of the eye.
Figures
References
-
- Bruzzone R, White TW, Paul DL. Connections with connexins: the molecular basis of direct intercellular signaling. Eur J Biochem. 1996;238:1–27. - PubMed
-
- Evans WH, Martin PE. Gap junctions: structure and function. Mol Membr Biol. 2002;19:121–136. - PubMed
-
- Willecke K, Eiberger J, Degen J, Eckardt D, Romualdi A, Guldenagel M, Deutsch U, Sohl G. Structural and functional diversity of connexin genes in the mouse and human genome. Biol Chem. 2002;383:725–737. - PubMed
-
- Gandolfi SA, Duncan G, Tomlinson J, Maraini G. Mammalian lens inter-fiber resistance is modulated by calcium and calmodulin. Curr Eye Res. 1990;9:533–541. - PubMed
-
- Crow JM, Atkinson MM, Johnson RG. Micromolar levels of intracellular calcium reduce gap junctional permeability in lens cultures. Invest Ophthalmol Visual Sci. 1994;35:3332–3341. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous