

Toxoplasma gondii sequesters centromeres to a specific nuclear region throughout the cell cycle
- PMID: 21321216
- PMCID: PMC3048097
- DOI: 10.1073/pnas.1006741108
Toxoplasma gondii sequesters centromeres to a specific nuclear region throughout the cell cycle
Abstract
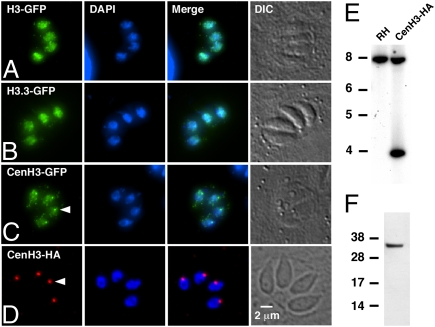
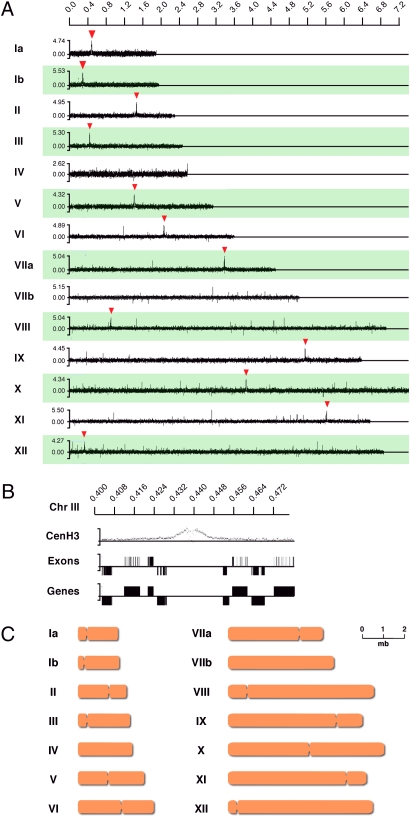
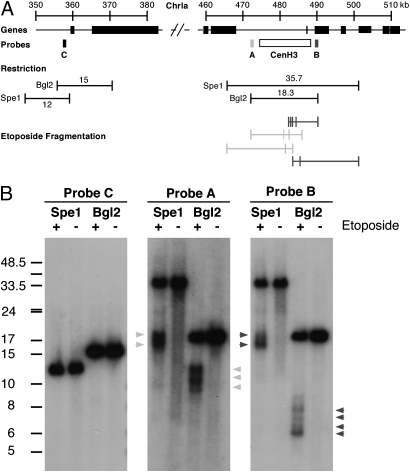
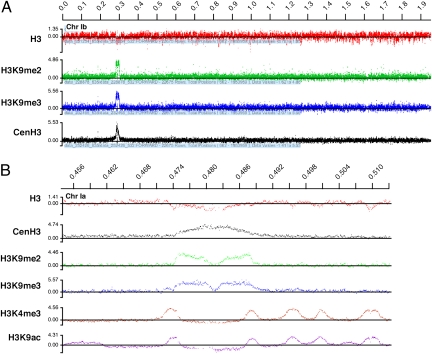
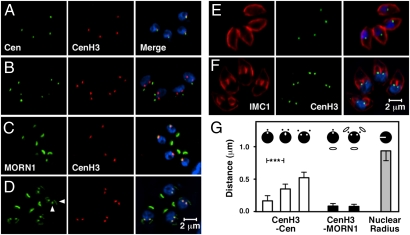
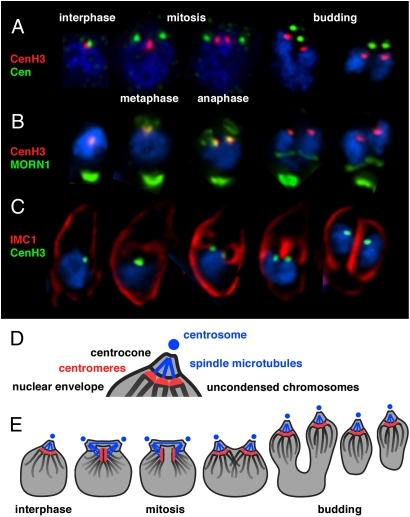
Members of the eukaryotic phylum Apicomplexa are the cause of important human diseases including malaria, toxoplasmosis, and cryptosporidiosis. These obligate intracellular parasites produce new invasive stages through a complex budding process. The budding cycle is remarkably flexible and can produce varied numbers of progeny to adapt to different host-cell niches. How this complex process is coordinated remains poorly understood. Using Toxoplasma gondii as a genetic model, we show that a key element to this coordination is the centrocone, a unique elaboration of the nuclear envelope that houses the mitotic spindle. Exploiting transgenic parasite lines expressing epitope-tagged centromeric H3 variant CenH3, we identify the centromeres of T. gondii chromosomes by hybridization of chromatin immunoprecipitations to genome-wide microarrays (ChIP-chip). We demonstrate that centromere attachment to the centrocone persists throughout the parasite cell cycle and that centromeres localize to a single apical region within the nucleus. Centromere sequestration provides a mechanism for the organization of the Toxoplasma nucleus and the maintenance of genome integrity.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Sibley LD. Intracellular parasite invasion strategies. Science. 2004;304:248–253. - PubMed
-
- Sheffield HG, Melton ML. The fine structure and reproduction of Toxoplasma gondii. J Parasitol. 1968;54:209–226. - PubMed
-
- Bannister LH, Hopkins JM, Fowler RE, Krishna S, Mitchell GH. A brief illustrated guide to the ultrastructure of Plasmodium falciparum asexual blood stages. Parasitol Today. 2000;16:427–433. - PubMed
-
- Speer CA, Dubey JP. Ultrastructure of shizonts and merozoites of Sarcocystis falcatula in the lungs of budgerigars (Melopsittacus undulatus) J Parasitol. 1999;85:630–637. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
