

Transport of receptors, receptor signaling complexes and ion channels via neuropeptide-secretory vesicles
- PMID: 21321602
- PMCID: PMC3203675
- DOI: 10.1038/cr.2011.29
Transport of receptors, receptor signaling complexes and ion channels via neuropeptide-secretory vesicles
Abstract
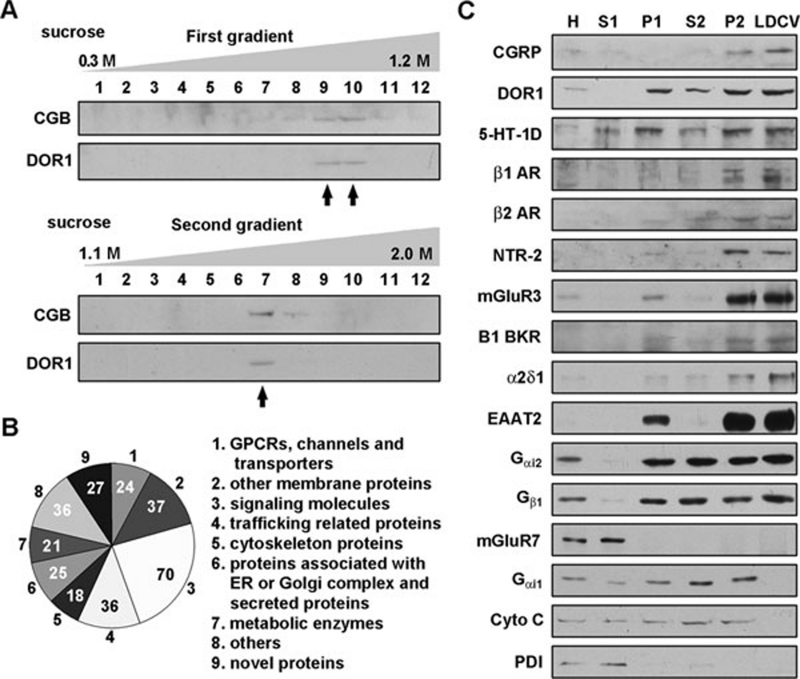
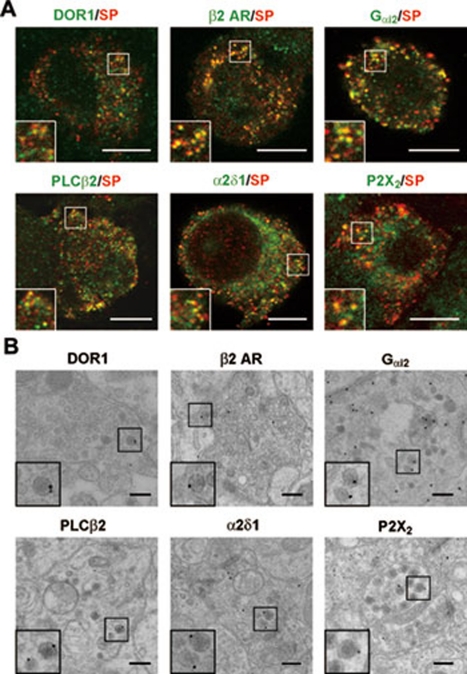
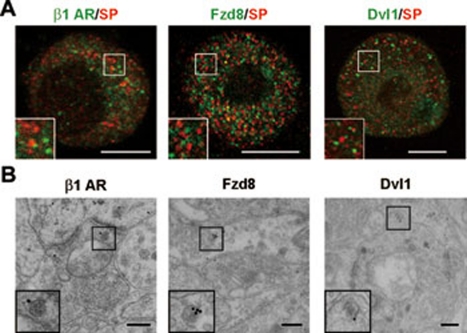
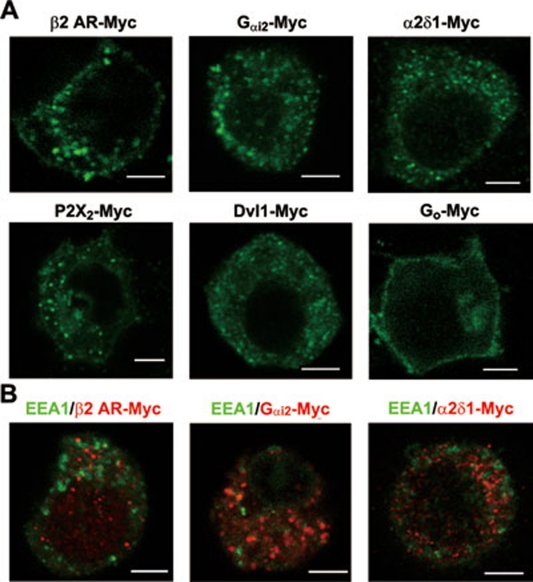
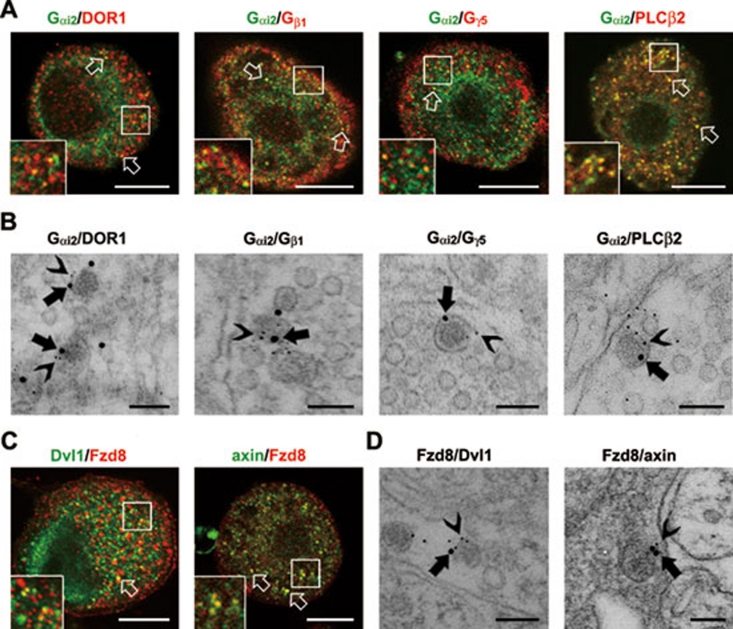
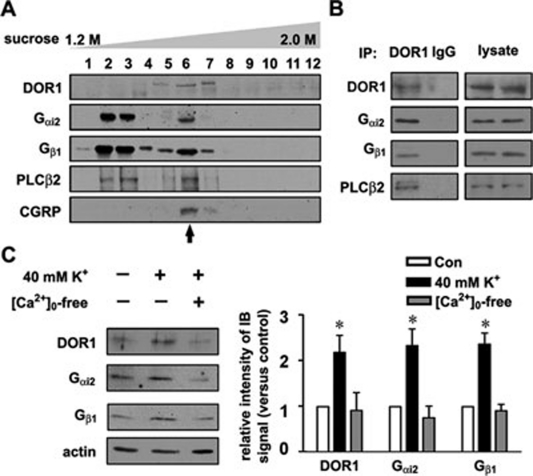
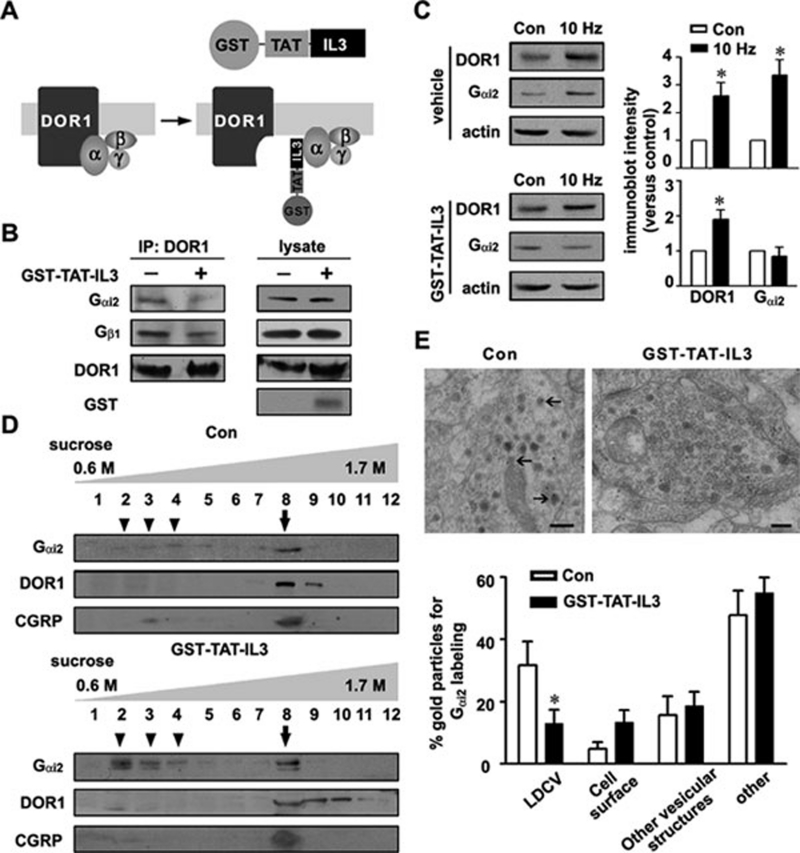
Stimulus-induced exocytosis of large dense-core vesicles (LDCVs) leads to discharge of neuropeptides and fusion of LDCV membranes with the plasma membrane. However, the contribution of LDCVs to the properties of the neuronal membrane remains largely unclear. The present study found that LDCVs were associated with multiple receptors, channels and signaling molecules, suggesting that neuronal sensitivity is modulated by an LDCV-mediated mechanism. Liquid chromatography-mass spectrometry combined with immunoblotting of subcellular fractions identified 298 proteins in LDCV membranes purified from the dorsal spinal cord, including G-protein-coupled receptors, G-proteins and other signaling molecules, ion channels and trafficking-related proteins. Morphological assays showed that δ-opioid receptor 1 (DOR1), β2 adrenergic receptor (AR), G(αi2), voltage-gated calcium channel α2δ1 subunit and P2X purinoceptor 2 were localized in substance P (SP)-positive LDCVs in small-diameter dorsal root ganglion neurons, whereas β1 AR, Wnt receptor frizzled 8 and dishevelled 1 were present in SP-negative LDCVs. Furthermore, DOR1/G(αi2)/G(β1γ5)/phospholipase C β2 complexes were associated with LDCVs. Blockade of the DOR1/G(αi2) interaction largely abolished the LDCV localization of G(αi2) and impaired stimulation-induced surface expression of G(αi2). Thus, LDCVs serve as carriers of receptors, ion channels and preassembled receptor signaling complexes, enabling a rapid, activity-dependent modulation of neuronal sensitivity.
Figures
References
-
- De Camilli P, Jahn R. Pathways to regulated exocytosis in neurons. Annu Rev Physiol. 1990;52:625–645. - PubMed
-
- Hökfelt T, Bartfai T, Bloom F. Neuropeptides: opportunities for drug discovery. Lancet Neurol. 2003;2:463–472. - PubMed
-
- Hannah MJ, Schmidt AA, Huttner WB. Synaptic vesicle biogenesis. Annu Rev Cell Dev Biol. 1999;15:733–798. - PubMed
-
- Takamori S, Holt M, Stenius K, et al. Molecular anatomy of a trafficking organelle. Cell. 2006;127:831–846. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
