

A meningococcal factor H binding protein mutant that eliminates factor H binding enhances protective antibody responses to vaccination
- PMID: 21325619
- PMCID: PMC3098282
- DOI: 10.4049/jimmunol.1003470
A meningococcal factor H binding protein mutant that eliminates factor H binding enhances protective antibody responses to vaccination
Abstract
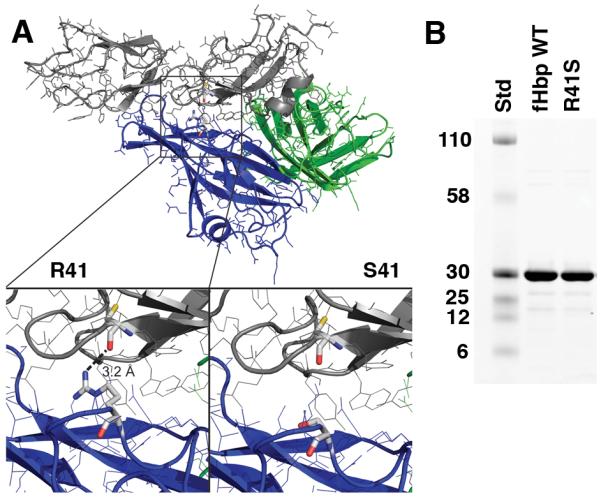
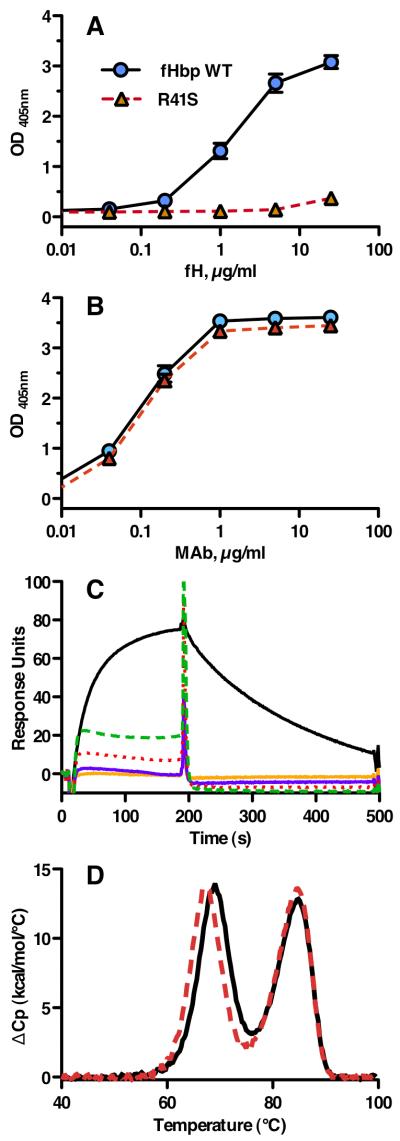
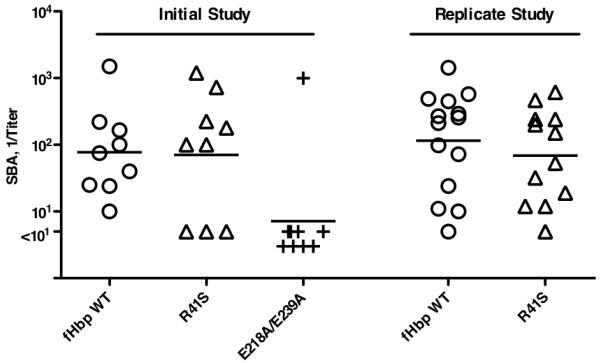
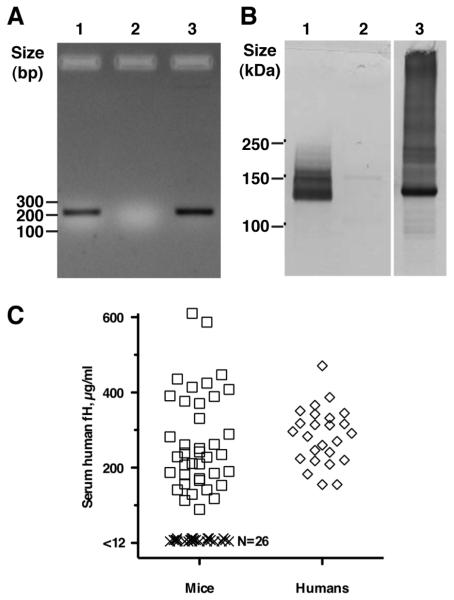
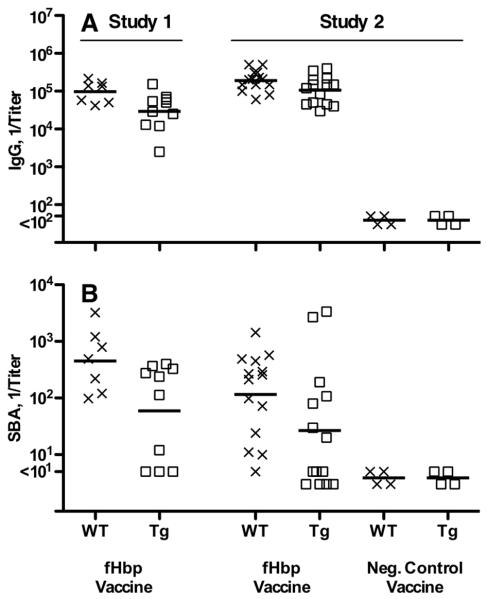
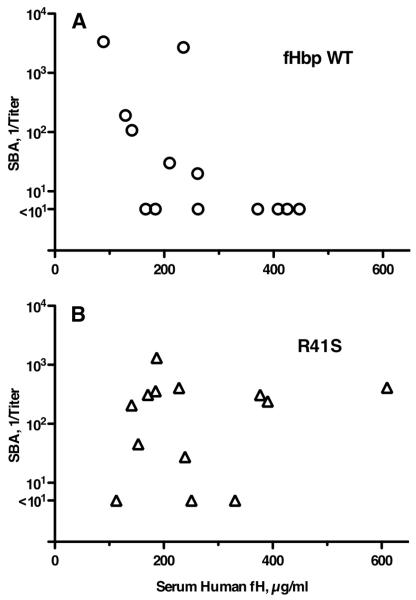
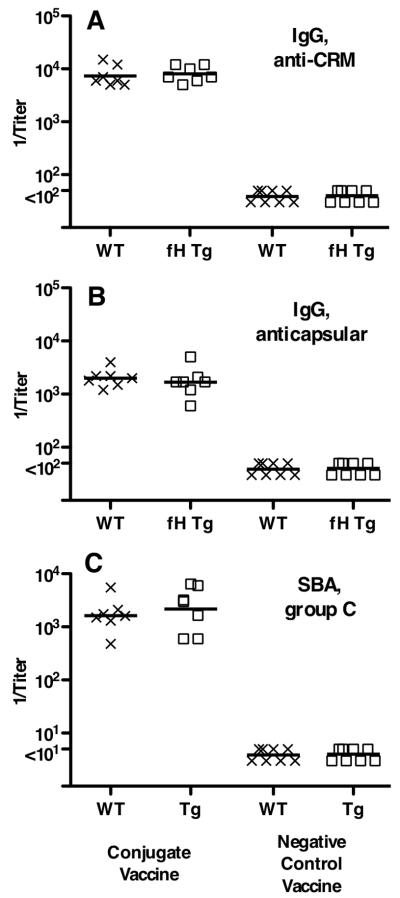
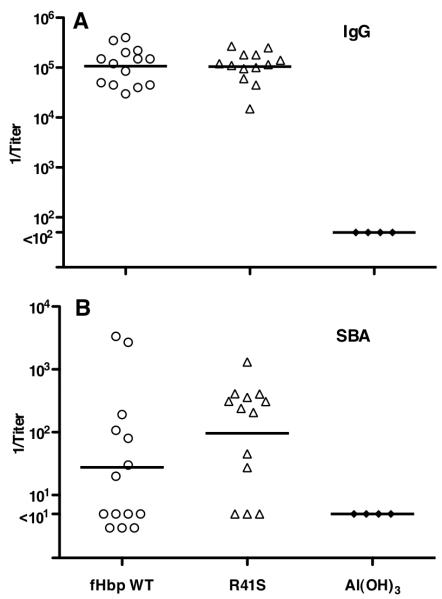
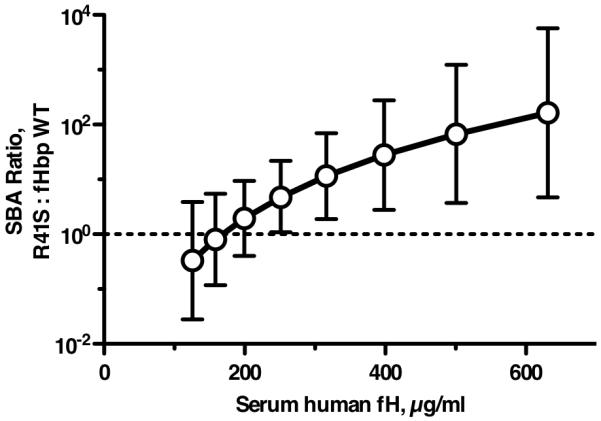
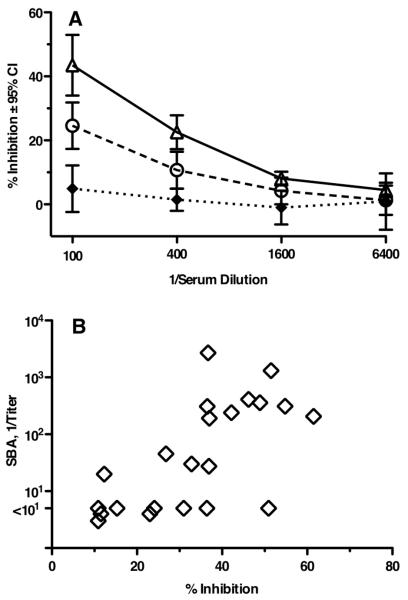
Certain pathogens recruit host complement inhibitors such as factor H (fH) to evade the immune system. Microbial complement inhibitor-binding molecules can be promising vaccine targets by eliciting Abs that neutralize this microbial defense mechanism. One such Ag, meningococcal factor H-binding protein (fHbp), was used in clinical trials before the protein was discovered to bind fH. The potential effect of fH binding on vaccine immunogenicity had not been assessed in experimental animals because fHbp binds human fH specifically. In this study, we developed a human fH transgenic mouse model. Transgenic mice immunized with fHbp vaccine had 4- to 8-fold lower serum bactericidal Ab responses than those of control mice whose native fH did not bind the vaccine. In contrast, Ab responses were unimpaired in transgenic mice immunized with a control meningococcal group C polysaccharide-protein conjugate vaccine. In transgenic mice, immunization with an fH nonbinding mutant of fHbp elicited Abs with higher bactericidal activity than that of fHbp vaccination itself. Abs elicited by the mutant fHbp more effectively blocked fH binding to wild-type fHbp than Abs elicited by fHbp that bound fH. Thus, a mutant fHbp vaccine that does not bind fH but that retains immunogenicity is predicted to be superior in humans to an fHbp vaccine that binds human fH. In the case of mutant fHbp vaccination, the resultant Ab responses may be directed more at epitopes in or near the fH binding site, which result in greater complement-mediated serum bactericidal activity; these epitopes may be obscured when human fH is bound to the wild-type fHbp vaccine.
Figures
References
-
- Serruto D, Rappuoli R, Scarselli M, Gros P, van Strijp JA. Molecular mechanisms of complement evasion: learning from staphylococci and meningococci. Nat Rev Microbiol. 2010;8:393–399. - PubMed
-
- Meri S, Jordens M, Jarva H. Microbial complement inhibitors as vaccines. Vaccine. 2008;26(Suppl 8):I113–117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- AI 082263/AI/NIAID NIH HHS/United States
- AI 084048/AI/NIAID NIH HHS/United States
- R37 AI032725/AI/NIAID NIH HHS/United States
- C06 RR 016226/RR/NCRR NIH HHS/United States
- R01 AI046464/AI/NIAID NIH HHS/United States
- U19 AI084048/AI/NIAID NIH HHS/United States
- R01 AI070955/AI/NIAID NIH HHS/United States
- AI 054544/AI/NIAID NIH HHS/United States
- R56 AI032725/AI/NIAID NIH HHS/United States
- AI 070955/AI/NIAID NIH HHS/United States
- AI 032725/AI/NIAID NIH HHS/United States
- R01 AI082263/AI/NIAID NIH HHS/United States
- C06 RR016226/RR/NCRR NIH HHS/United States
- R01 AI032725/AI/NIAID NIH HHS/United States
- R01 AI 046464/AI/NIAID NIH HHS/United States
- R01 AI054544/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
