

Action understanding and active inference
- PMID: 21327826
- PMCID: PMC3491875
- DOI: 10.1007/s00422-011-0424-z
Action understanding and active inference
Abstract
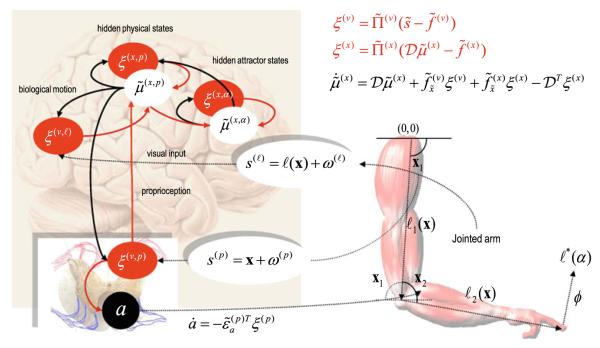
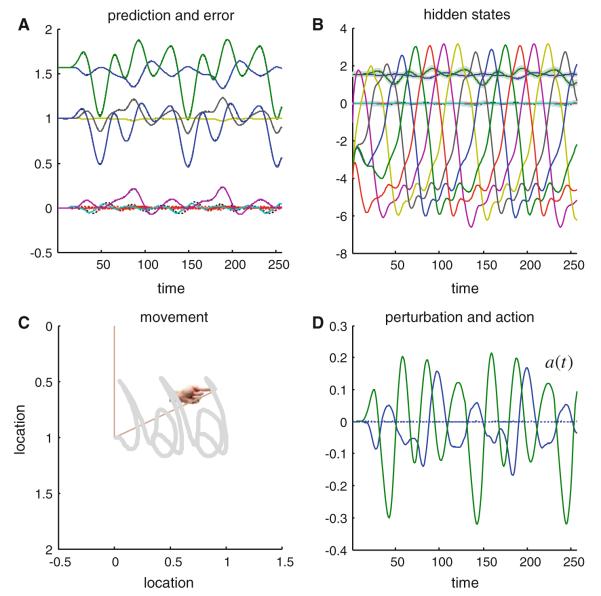
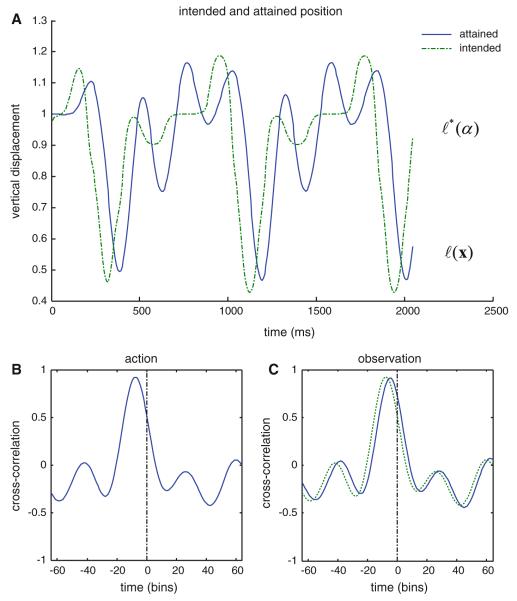
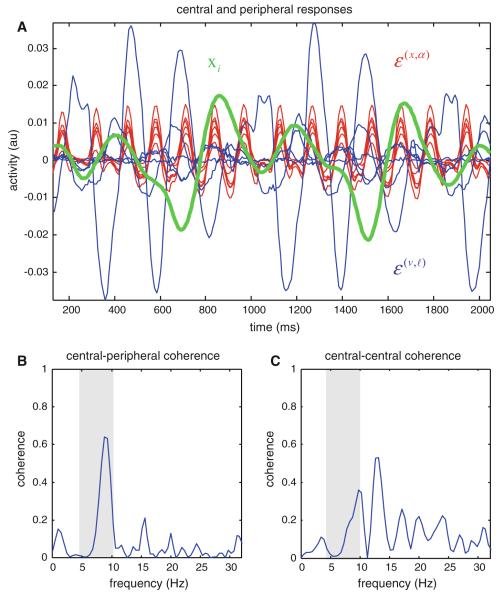
We have suggested that the mirror-neuron system might be usefully understood as implementing Bayes-optimal perception of actions emitted by oneself or others. To substantiate this claim, we present neuronal simulations that show the same representations can prescribe motor behavior and encode motor intentions during action-observation. These simulations are based on the free-energy formulation of active inference, which is formally related to predictive coding. In this scheme, (generalised) states of the world are represented as trajectories. When these states include motor trajectories they implicitly entail intentions (future motor states). Optimizing the representation of these intentions enables predictive coding in a prospective sense. Crucially, the same generative models used to make predictions can be deployed to predict the actions of self or others by simply changing the bias or precision (i.e. attention) afforded to proprioceptive signals. We illustrate these points using simulations of handwriting to illustrate neuronally plausible generation and recognition of itinerant (wandering) motor trajectories. We then use the same simulations to produce synthetic electrophysiological responses to violations of intentional expectations. Our results affirm that a Bayes-optimal approach provides a principled framework, which accommodates current thinking about the mirror-neuron system. Furthermore, it endorses the general formulation of action as active inference.
Figures
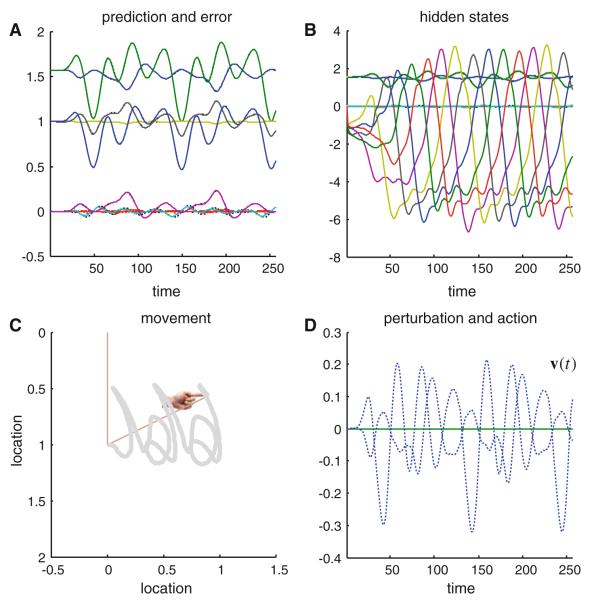

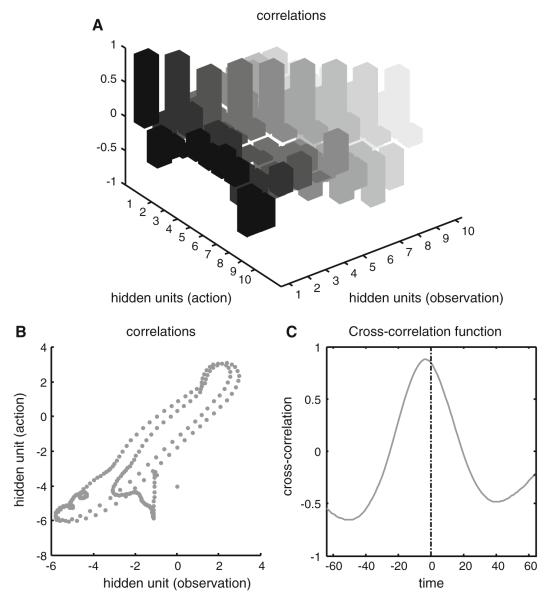
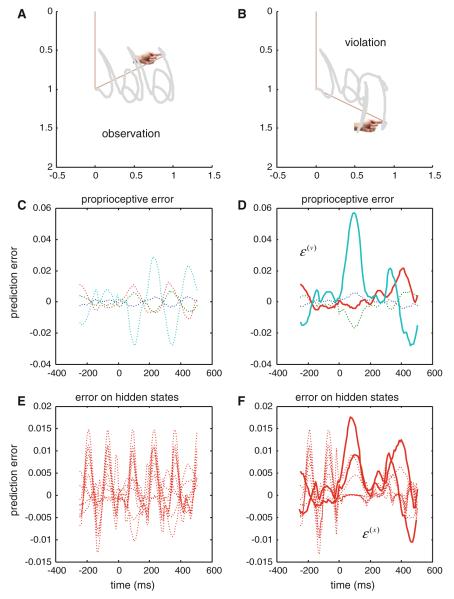
References
-
- Afraimovich V, Tristan I, Huerta R, Rabinovich MI. Winnerless competition principle and prediction of the transient dynamics in a Lotka–Volterra model. Chaos. 2008;18(4):043103. - PubMed
-
- Allison T, Puce A, McCarthy G. Social perception from visual cues: role of the STS region. Trends Cogn Sci. 2000;4:267–278. - PubMed
-
- Arbib MA. From grasp to language: embodied concepts and the challenge of abstraction. J Physiol (Paris) 2008;102(1-3):4–20. - PubMed
-
- Arbib MA. Mirror system activity for action and language is embedded in the integration of dorsal and ventral pathways. Brain Lang. 2010;112(1):12–24. - PubMed
-
- Ballard DH, Hinton GE, Sejnowski TJ. Parallel visual computation. Nature. 1983;306:21–26. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
