

A nanomolar-potency small molecule inhibitor of regulator of G-protein signaling proteins
- PMID: 21329361
- PMCID: PMC3090679
- DOI: 10.1021/bi1019622
A nanomolar-potency small molecule inhibitor of regulator of G-protein signaling proteins
Abstract
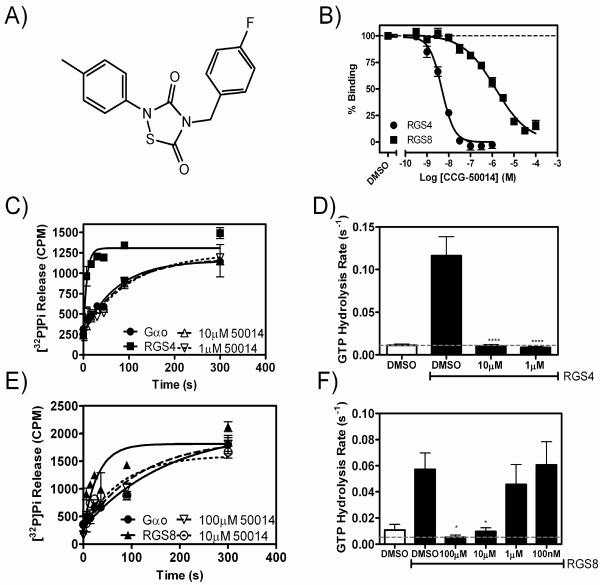
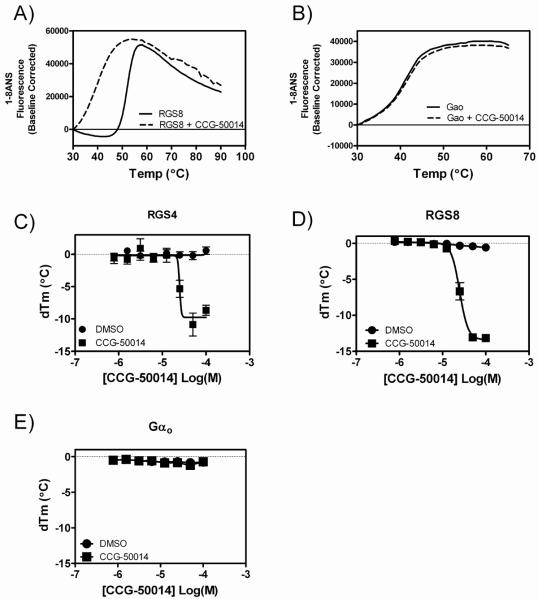
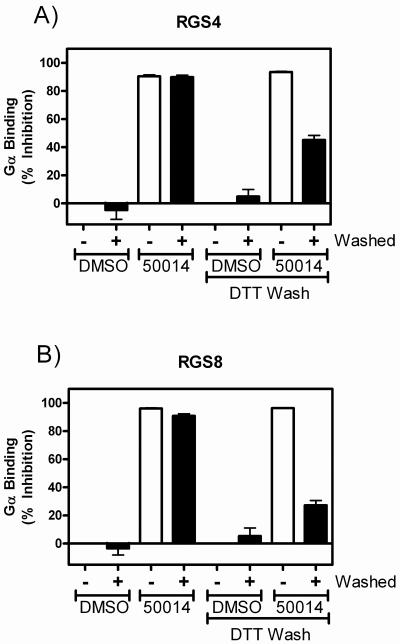
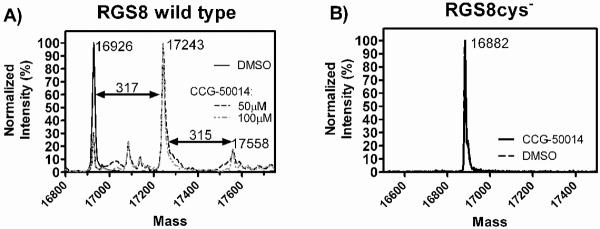
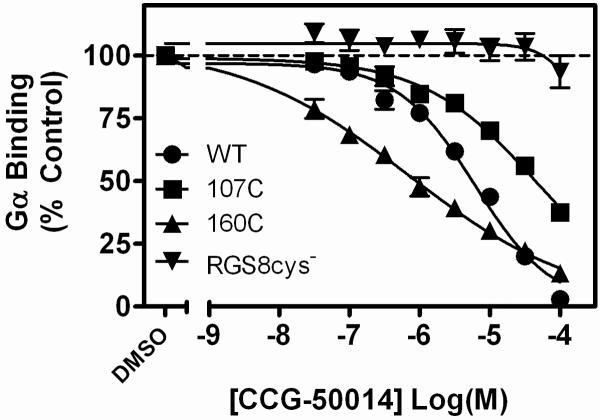
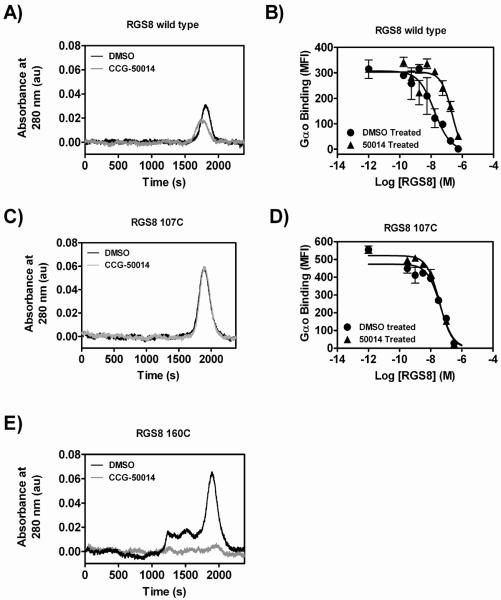
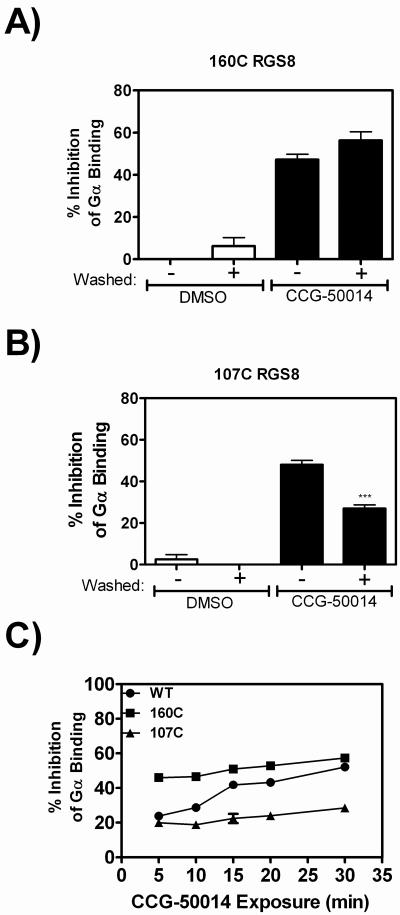
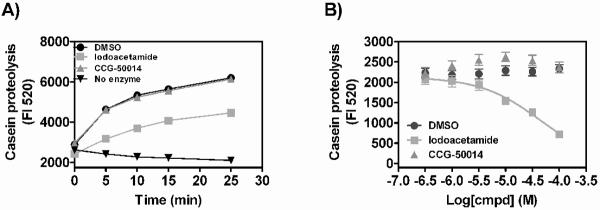
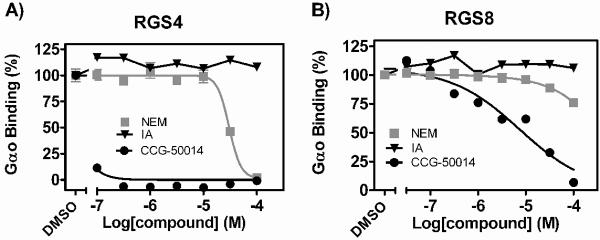
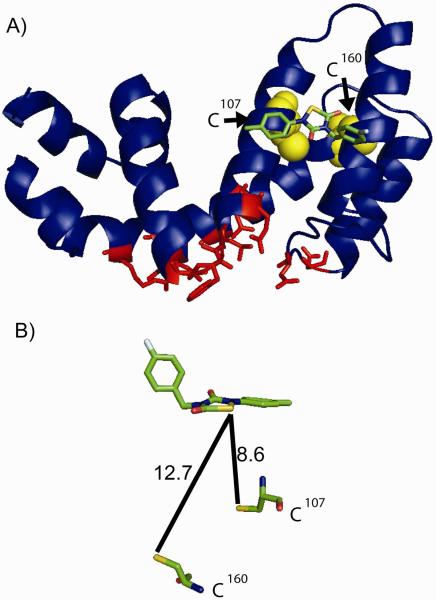
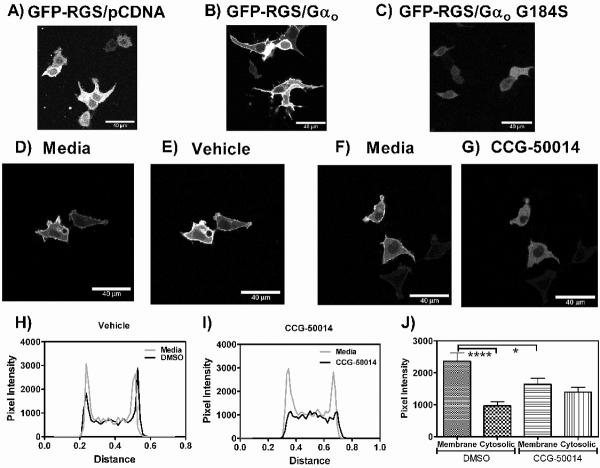
Regulators of G-protein signaling (RGS) proteins are potent negative modulators of signal transduction through G-protein-coupled receptors. They function by binding to activated (GTP-bound) Gα subunits and accelerating the rate of GTP hydrolysis. Modulation of RGS activity by small molecules is an attractive mechanism for fine-tuning GPCR signaling for therapeutic and research purposes. Here we describe the pharmacologic properties and mechanism of action of CCG-50014, the most potent small molecule RGS inhibitor to date. It has an IC(50) for RGS4 of 30 nM and is >20-fold selective for RGS4 over other RGS proteins. CCG-50014 binds covalently to the RGS, forming an adduct on two cysteine residues located in an allosteric regulatory site. It is not a general cysteine alkylator as it does not inhibit activity of the cysteine protease papain at concentrations >3000-fold higher than those required to inhibit RGS4 function. It is also >1000-fold more potent as an RGS4 inhibitor than are the cysteine alkylators N-ethylmaleimide and iodoacetamide. Analysis of the cysteine reactivity of the compound shows that compound binding to Cys(107) in RGS8 inhibits Gα binding in a manner that can be reversed by cleavage of the compound-RGS disulfide bond. If the compound reacts with Cys(160) in RGS8, the adduct induces RGS denaturation, and activity cannot be restored by removal of the compound. The high potency and good selectivity of CCG-50014 make it a useful tool for studying the functional roles of RGS4.
Figures
References
-
- Lewis JA, Lebois EP, Lindsley CW. Allosteric modulation of kinases and GPCRs: design principles and structural diversity. Curr Opin Chem Biol. 2008;12:269–280. - PubMed
-
- Blazer LL, Neubig RR. Small molecule protein-protein interaction inhibitors as CNS therapeutic agents: current progress and future hurdles. Neuropsychopharmacology. 2009;34:126–141. - PubMed
-
- Berman DM, Kozasa T, Gilman AG. The GTPase-activating protein RGS4 stabilizes the transition state for nucleotide hydrolysis. J Biol Chem. 1996;271:27209–27212. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
