

Purinergic mechanisms in gliovascular coupling
- PMID: 21329762
- PMCID: PMC3070818
- DOI: 10.1016/j.semcdb.2011.02.010
Purinergic mechanisms in gliovascular coupling
Abstract
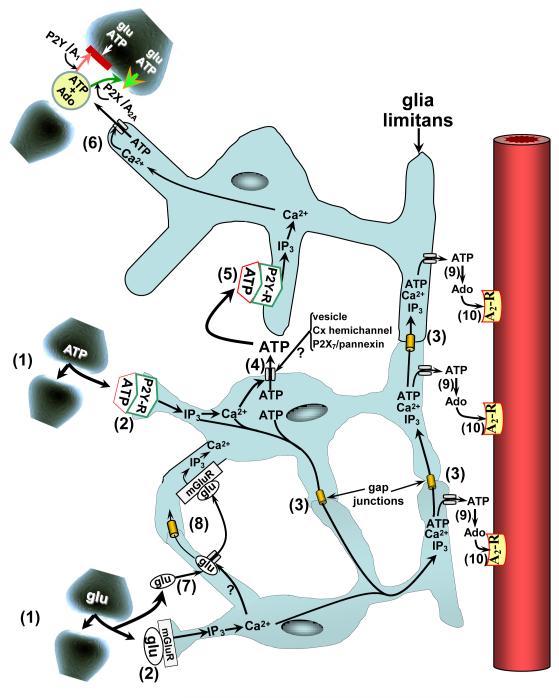
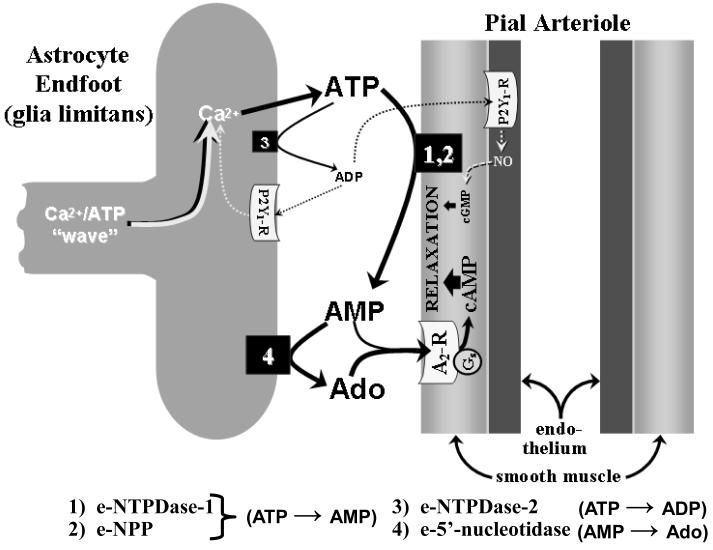
Regional elevations in cerebral blood flow (CBF) often occur in response to localized increases in cerebral neuronal activity. An ever expanding literature has linked this neurovascular coupling process to specific signaling pathways involving neuronal synapses, astrocytes and cerebral arteries and arterioles. Collectively, these structures are termed the "neurovascular unit" (NVU). Astrocytes are thought to be the cornerstone of the NVU. Thus, not only do astrocytes "detect" increased synaptic activity, they can transmit that information to proximal and remote astrocytic sites often through a Ca(2+)- and ATP-related signaling process. At the vascular end of the NVU, a Ca(2+)-dependent formation and release of vasodilators, or substances linked to vasodilation, can occur. The latter category includes ATP, which upon its appearance in the extracellular compartment, can be rapidly converted to the potent vasodilator, adenosine, via the action of ecto-nucleotidases. In the present review, we give consideration to experimental model-specific variations in purinergic influences on gliovascular signaling mechanisms, focusing on the cerebral cortex. In that discussion, we compare findings obtained using in vitro (rodent brain slice) models and multiple in vivo models (2-photon imaging; somatosensory stimulation-evoked cortical hyperemia; and sciatic nerve stimulation-evoked pial arteriolar dilation). Additional attention is given to the importance of upstream (remote) vasodilation; the key role played by extracellular ATP hydrolysis (via ecto-nucleotidases) in gliovascular coupling; and interactions among multiple signaling pathways.
Copyright © 2011 Elsevier Ltd. All rights reserved.
Figures
References
-
- Iadecola C, Nedergaard M. Glial regulation of the cerebral microvasculature. Nat Neurosci. 2007;10:1369–76. - PubMed
-
- Koehler RC, Roman RJ, Harder DR. Astrocytes and the regulation of cerebral blood flow. Trends Neurosci. 2009;32:160–9. - PubMed
-
- Haydon PG, Carmignoto G. Astrocyte control of synaptic transmission and neurovascular coupling. Physiol Rev. 2006;86:1009–31. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
