

The dlt operon confers resistance to cationic antimicrobial peptides in Clostridium difficile
- PMID: 21330441
- PMCID: PMC3140582
- DOI: 10.1099/mic.0.045997-0
The dlt operon confers resistance to cationic antimicrobial peptides in Clostridium difficile
Abstract
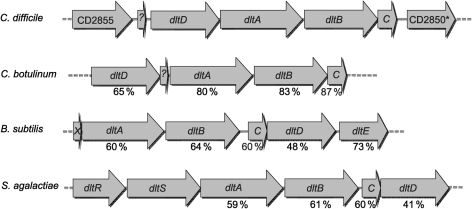
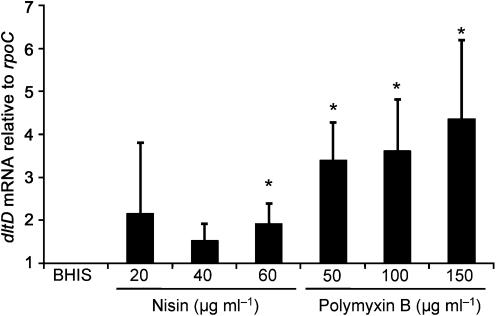
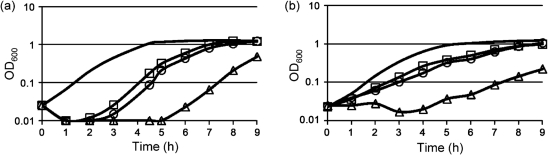
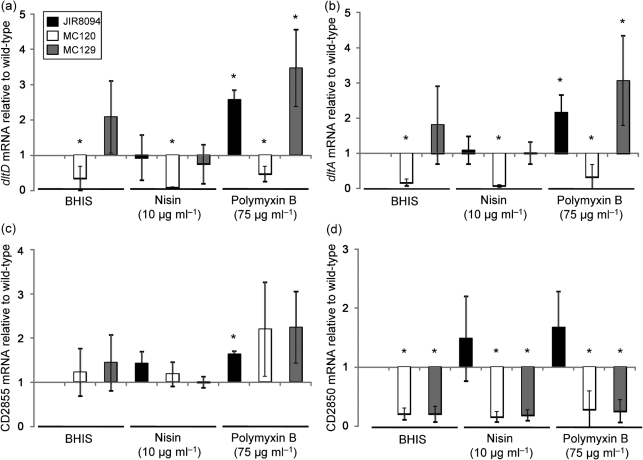
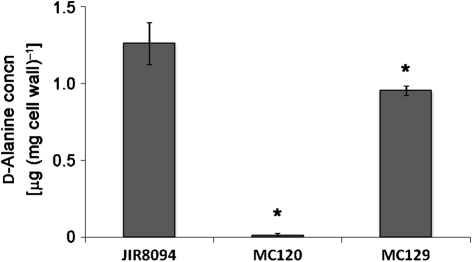
The dlt operon in Gram-positive bacteria encodes proteins that are necessary for the addition of d-alanine to teichoic acids of the cell wall. The addition of d-alanine to the cell wall results in a net positive charge on the bacterial cell surface and, as a consequence, can decrease the effectiveness of antimicrobials, such as cationic antimicrobial peptides (CAMPs). Although the roles of the dlt genes have been studied for some Gram-positive organisms, the arrangement of these genes in Clostridium difficile and the life cycle of the bacterium in the host are markedly different from those of other pathogens. In the current work, we determined the contribution of the putative C. difficile dlt operon to CAMP resistance. Our data indicate that the dlt operon is necessary for full resistance of C. difficile to nisin, gallidermin, polymyxin B and vancomycin. We propose that the d-alanylation of teichoic acids provides protection against antimicrobial peptides that may be essential for growth of C. difficile in the host.
Figures
References
-
- Abi Khattar Z., Rejasse A., Destoumieux-Garzón D., Escoubas J. M., Sanchis V., Lereclus D., Givaudan A., Kallassy M., Nielsen-Leroux C., Gaudriault S. (2009). The dlt operon of Bacillus cereus is required for resistance to cationic antimicrobial peptides and for virulence in insects. J Bacteriol 191, 7063–7073. 10.1128/JB.00892-09 - DOI - PMC - PubMed
-
- Allen, D. (2008) Transcriptional regulation of the dlt operon in Enterococcus faecalis and further characterization of a dltA mutant. MS thesis, Kansas State University, Manhattan, KS.
-
- Bartlett J. G., Onderdonk A. B., Cisneros R. L., Kasper D. L. (1977). Clindamycin-associated colitis due to a toxin-producing species of Clostridium in hamsters. J Infect Dis 136, 701–705. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases