

Review
doi: 10.1007/s10577-011-9191-x.
Springs, clutches and motors: driving forward kinetochore mechanism by modelling
Affiliations
- PMID: 21331796
- PMCID: PMC3078324
- DOI: 10.1007/s10577-011-9191-x
Item in Clipboard
Review
Springs, clutches and motors: driving forward kinetochore mechanism by modelling
Chromosome Res.
2011 Apr.
Abstract
As a mechanical system, the kinetochore can be viewed as a set of interacting springs, clutches and motors; the problem of kinetochore mechanism is now one of understanding how these functional modules assemble, disassemble and interact with one another to give rise to the emergent properties of the system. The sheer complexity of the kinetochore system points to a future requirement for data-driven mathematical modelling and statistical analysis based on quantitative empirical measurement of sister kinetochore trajectories. Here, we review existing models of chromosome motion in the context of recent advances in our understanding of kinetochore molecular biology.
Figures
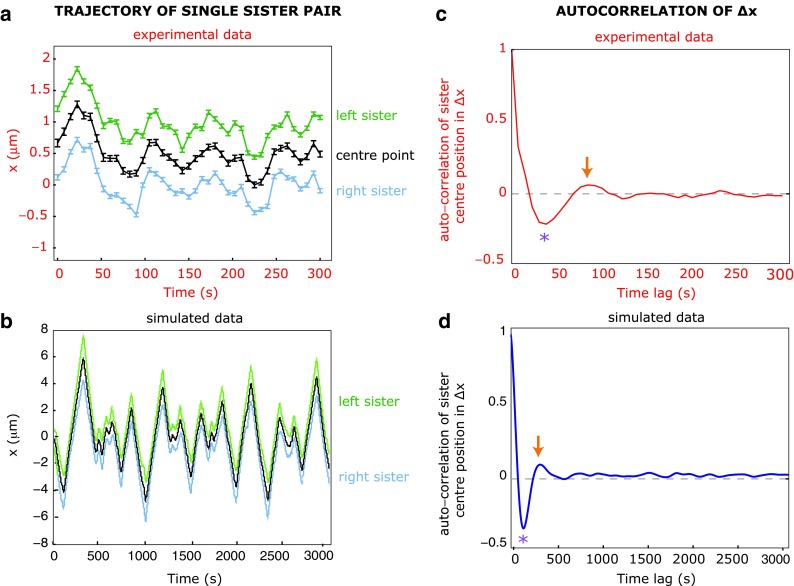
Comparison of experimental and simulated data of sister kinetochore oscillations in metaphase. a Trajectory of a single sister kinetochore pair undergoing oscillations along the normal (x) to the metaphase plate. Coordinate positions of right (blue line) and left (green line) sisters plus the centre point between them (black line) are from experimental data of kinetochores in human HeLa cells (Vladimirou and McAinsh, unpublished data). b Trajectory of a single sister kinetochore pair undergoing oscillations in metaphase. Data was simulated using the mathematical model from Joglekar and Hunt , which is largely based on parameters from PTK-1 cells, hence the longer timescale for oscillations compared to human cells. c, d Autocorrelation of the centre positions of sister pairs from experimental (50 trajectories; red line; c) and simulated data (100 trajectories; blue line; d). The half period (purple star) and full period (orange arrow) of the oscillation in the autocorrelation curves are indicated
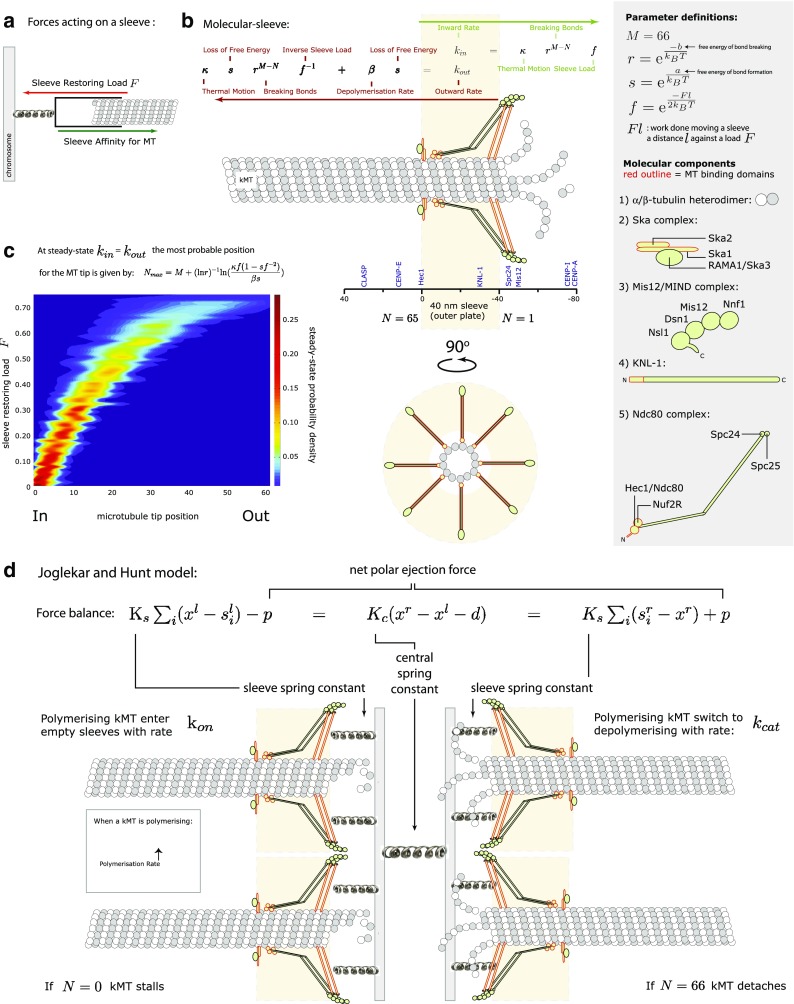
Hill sleeve and Joglekar and Hunt model. a A Hill sleeve balances its affinity for a MT (green arrow) against the restoring load (red arrow) pulling it back to the chromosome. b Contemporary view (upper panel from the side; lower panel is a 90° rotation showing end-on view) of a Hill sleeve. Binding sites inside the sleeve are shown as KMN networks and Ska complexes. These form multiple attachment sites for the MT as it penetrates into the sleeve, which is the outer plate of the kinetochore. The inward (green arrow) and outward (red arrow) movement rates of the MT are shown and depend on the position of the MT inside the sleeve and the restoring load on the sleeve. Position of kinetochore components within the kinetochore is based on super-resolution imaging and adapted from Wan et al. . Subunit composition and organisation of complexes is based on Cheeseman et al. ; Maskell et al. ; Petrovic et al. and Wan et al. . c A Monte Carlo simulation of the kMT tip position inside the sleeve gives the probability density distribution over position and load (based on rate equations from Joglekar and Hunt 2002). Higher loads on the sleeve increase the probability of MT tip shifting to positions further and further out of the sleeve. d The Joglekar and Hunt model is based on a force balance between the kinetochores, sleeves and polar ejection forces. The positions of the sleeves are determined by the kinetics of the sleeve-bound MT as determined by Hill’s rate equations (Hill 1985). Polymerising MTs enter empty sleeves with a rate k
on and switch to depolymerising with a rate k
cat. N is the position of the MT tip inside the sleeve. When N = 1 the MT is fully inserted, whereas the MT detaches from the sleeve when N = 66
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
