

The karyopherin Kap95 and the C-termini of Rfa1, Rfa2, and Rfa3 are necessary for efficient nuclear import of functional RPA complex proteins in Saccharomyces cerevisiae
- PMID: 21332387
- PMCID: PMC3168979
- DOI: 10.1089/dna.2010.1071
The karyopherin Kap95 and the C-termini of Rfa1, Rfa2, and Rfa3 are necessary for efficient nuclear import of functional RPA complex proteins in Saccharomyces cerevisiae
Abstract
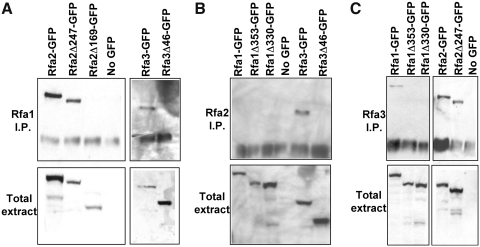
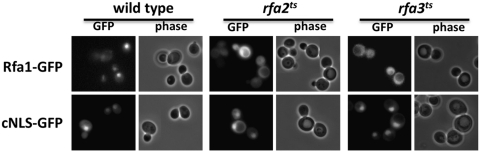
Nuclear protein import in eukaryotic cells is mediated by karyopherin proteins, which bind to specific nuclear localization signals on substrate proteins and transport them across the nuclear envelope and into the nucleus. Replication protein A (RPA) is a nuclear protein comprised of three subunits (termed Rfa1, Rfa2, and Rfa3 in Saccharomyces cerevisiae) that binds single-stranded DNA and is essential for DNA replication, recombination, and repair. RPA associates with two different karyopherins in yeast, Kap95, and Msn5/Kap142. However, it is unclear which of these karyopherins is responsible for RPA nuclear import. We have generated GFP fusion proteins with each of the RPA subunits and demonstrate that these Rfa-GFP chimeras are functional in yeast cells. The intracellular localization of the RPA proteins in live cells is similar in wild-type and msn5Δ deletion strains but becomes primarily cytoplasmic in cells lacking functional Kap95. Truncating the C-terminus of any of the RPA subunits results in mislocalization of the proteins to the cytoplasm and a loss of protein-protein interactions between the subunits. Our data indicate that Kap95 is likely the primary karyopherin responsible for RPA nuclear import in yeast and that the C-terminal regions of Rfa1, Rfa2, and Rfa3 are essential for efficient nucleocytoplasmic transport of each RPA subunit.
Figures
Similar articles
-
The karyopherin Msn5/Kap142 requires Nup82 for nuclear export and performs a function distinct from translocation in RPA protein import.J Biol Chem. 2004 Oct 15;279(42):43530-9. doi: 10.1074/jbc.M407641200. Epub 2004 Aug 4. J Biol Chem. 2004. PMID: 15294903
-
The karyopherin Kap142p/Msn5p mediates nuclear import and nuclear export of different cargo proteins.J Cell Biol. 2001 Feb 19;152(4):729-40. doi: 10.1083/jcb.152.4.729. J Cell Biol. 2001. PMID: 11266464 Free PMC article.
-
Yeast karyopherin Kap95 is required for cell cycle progression at Start.BMC Cell Biol. 2010 Jun 29;11:47. doi: 10.1186/1471-2121-11-47. BMC Cell Biol. 2010. PMID: 20587033 Free PMC article.
-
Nuclear import by karyopherin-βs: recognition and inhibition.Biochim Biophys Acta. 2011 Sep;1813(9):1593-606. doi: 10.1016/j.bbamcr.2010.10.014. Epub 2010 Oct 26. Biochim Biophys Acta. 2011. PMID: 21029754 Free PMC article. Review.
-
Molecular mechanisms of nuclear protein transport.Crit Rev Eukaryot Gene Expr. 1997;7(1-2):61-72. doi: 10.1615/critreveukargeneexpr.v7.i1-2.40. Crit Rev Eukaryot Gene Expr. 1997. PMID: 9034715 Review.
Cited by
-
Replication protein A: a multifunctional protein with roles in DNA replication, repair and beyond.NAR Cancer. 2020 Sep 25;2(3):zcaa022. doi: 10.1093/narcan/zcaa022. eCollection 2020 Sep. NAR Cancer. 2020. PMID: 34316690 Free PMC article.
-
Chaperoning RPA during DNA metabolism.Curr Genet. 2019 Aug;65(4):857-864. doi: 10.1007/s00294-019-00945-3. Epub 2019 Feb 22. Curr Genet. 2019. PMID: 30796471 Review.
-
Karyopherin-mediated nucleocytoplasmic transport.Nat Rev Mol Cell Biol. 2022 May;23(5):307-328. doi: 10.1038/s41580-021-00446-7. Epub 2022 Jan 20. Nat Rev Mol Cell Biol. 2022. PMID: 35058649 Free PMC article. Review.
-
Yeast importin-β is required for nuclear import of the Mig2 repressor.BMC Cell Biol. 2012 Nov 6;13:31. doi: 10.1186/1471-2121-13-31. BMC Cell Biol. 2012. PMID: 23131016 Free PMC article.
-
Microscopy analysis of the smallest subunit of the RPA complex, Rfa3p, prompts consideration of how RPA subunits gather at single-stranded DNA sites.MicroPubl Biol. 2021 Oct 27;2021:10.17912/micropub.biology.000493. doi: 10.17912/micropub.biology.000493. eCollection 2021. MicroPubl Biol. 2021. PMID: 34723142 Free PMC article.
References
-
- Adams A. Gottschling D.E. Kaiser C.A. Stearns T. Cold Spring Harbor Laboratory Press; Cold Spring Harbor, NY: 1997. Methods in Yeast Genetics.
-
- Bakhrat A. Baranes-Bachar K. Reshef D. Voloshin O. Krichevsky O. Raveh D. Nuclear export of Ho endonuclease of yeast via Msn5. Curr Genet. 2008;54:271–281. - PubMed
-
- Belanger K.D. Simmons L.A. Roth J.K. VanderPloeg K.A. Lichten L.B. Fahrenkrog B. The karyopherin Msn5/Kap142 requires Nup82 for nuclear export and performs a function distinct from translocation in RPA protein import. J Biol Chem. 2004;279:43530–43539. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
