

T cells fail to develop in the human skin-cell explants system; an inconvenient truth
- PMID: 21332988
- PMCID: PMC3056828
- DOI: 10.1186/1471-2172-12-17
T cells fail to develop in the human skin-cell explants system; an inconvenient truth
Abstract
Background: Haplo-identical hematopoietic stem cell (HSC) transplantation is very successful in eradicating haematological tumours, but the long post-transplant T-lymphopenic phase is responsible for high morbidity and mortality rates. Clark et al. have described a skin-explant system capable of producing host-tolerant donor-HSC derived T-cells. Because this T-cell production platform has the potential to replenish the T-cell levels following transplantation, we set out to validate the skin-explant system.
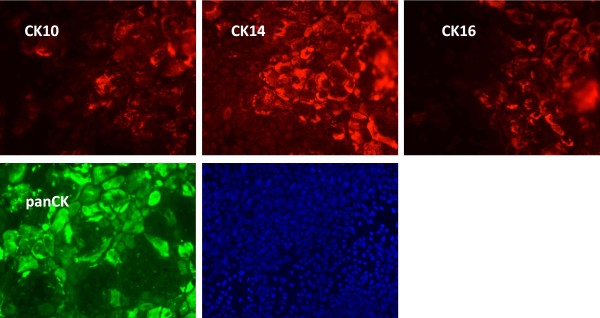
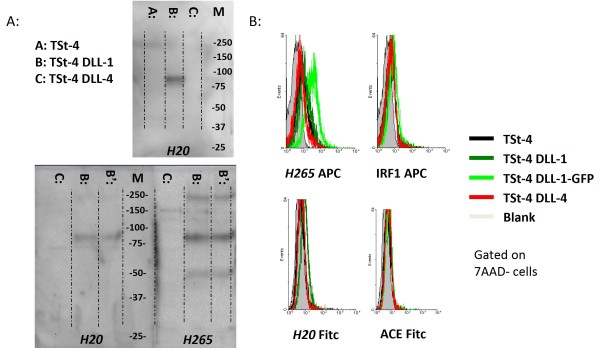
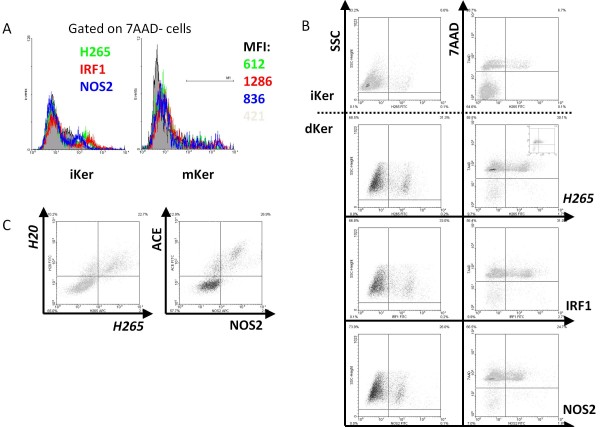
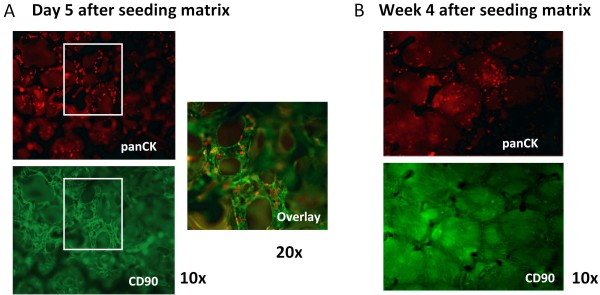
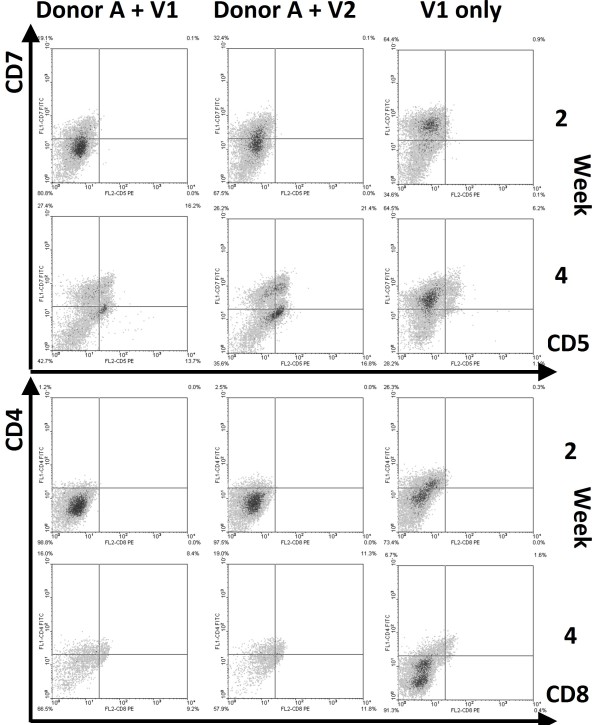
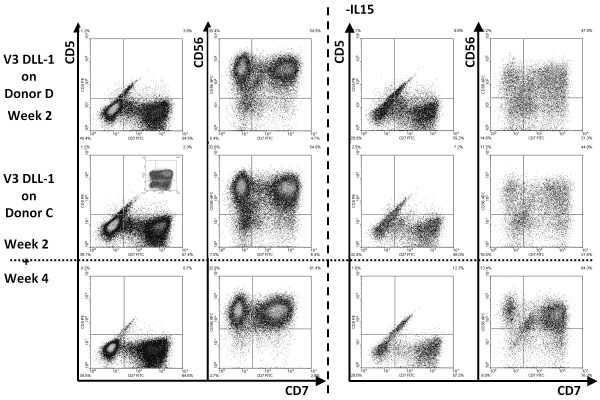
Results: Following the published procedures, while using the same commercial components, it was impossible to reproduce the skin-explant conditions required for HSC differentiation towards mature T-cells. The keratinocyte maturation procedure resulted in fragile cells with minimum expression of delta-like ligand (DLL). In most experiments the generated cells failed to adhere to carriers or were quickly outcompeted by fibroblasts. Consequently it was not possible to reproduce cell-culture conditions required for HSC differentiation into functional T-cells. Using cell-lines over-expressing DLL, we showed that the antibodies used by Clark et al. were unable to detect native DLL, but instead stained 7AAD+ cells. Therefore, it is unlikely that the observed T-lineage commitment from HSC is mediated by DLL expressed on keratinocytes. In addition, we did confirm expression of the Notch-ligand Jagged-1 by keratinocytes.
Conclusions: Currently, and unfortunately, it remains difficult to explain the development or growth of T-cells described by Clark et al., but for the fate of patients suffering from lymphopenia it is essential to both reproduce and understand how these co-cultures really "work". Fortunately, alternative procedures to speed-up T-cell reconstitution are being established and validated and may become available for patients in the near future.
Figures

Similar articles
-
Direct comparison of Dll1- and Dll4-mediated Notch activation levels shows differential lymphomyeloid lineage commitment outcomes.J Immunol. 2010 Jul 15;185(2):867-76. doi: 10.4049/jimmunol.1000782. Epub 2010 Jun 14. J Immunol. 2010. PMID: 20548034
-
Induction of T Cell Development In Vitro by Delta-Like (Dll)-Expressing Stromal Cells.Methods Mol Biol. 2016;1323:159-67. doi: 10.1007/978-1-4939-2809-5_14. Methods Mol Biol. 2016. PMID: 26294407
-
T-lymphoid differentiation potential measured in vitro is higher in CD34+CD38-/lo hematopoietic stem cells from umbilical cord blood than from bone marrow and is an intrinsic property of the cells.Haematologica. 2011 May;96(5):646-54. doi: 10.3324/haematol.2010.036343. Epub 2011 Feb 17. Haematologica. 2011. PMID: 21330325 Free PMC article.
-
T-cell development, doing it in a dish.Immunol Rev. 2006 Feb;209:95-102. doi: 10.1111/j.0105-2896.2006.00353.x. Immunol Rev. 2006. PMID: 16448536 Review.
-
Regulation of intrathymic T-cell development by Lunatic Fringe- Notch1 interactions.Immunol Rev. 2006 Feb;209:76-94. doi: 10.1111/j.0105-2896.2006.00360.x. Immunol Rev. 2006. PMID: 16448535 Review.
Cited by
-
Engineering approaches for regeneration of T lymphopoiesis.Biomater Res. 2016 Jun 29;20:20. doi: 10.1186/s40824-016-0067-1. eCollection 2016. Biomater Res. 2016. PMID: 27358746 Free PMC article. Review.
-
Engineered T cells from induced pluripotent stem cells: from research towards clinical implementation.Front Immunol. 2024 Jan 12;14:1325209. doi: 10.3389/fimmu.2023.1325209. eCollection 2023. Front Immunol. 2024. PMID: 38283344 Free PMC article. Review.
-
Novel immortal human cell lines reveal subpopulations in the nucleus pulposus.Arthritis Res Ther. 2014 Jun 27;16(3):R135. doi: 10.1186/ar4597. Arthritis Res Ther. 2014. PMID: 24972717 Free PMC article.
-
A simple model system enabling human CD34(+) cells to undertake differentiation towards T cells.PLoS One. 2013 Jul 23;8(7):e69572. doi: 10.1371/journal.pone.0069572. Print 2013. PLoS One. 2013. PMID: 23894504 Free PMC article.
-
In Vitro T-Cell Generation From Adult, Embryonic, and Induced Pluripotent Stem Cells: Many Roads to One Destination.Stem Cells. 2015 Nov;33(11):3174-80. doi: 10.1002/stem.2115. Epub 2015 Aug 11. Stem Cells. 2015. PMID: 26227158 Free PMC article. Review.
References
-
- Mackall CL, Stein D, Fleisher TA, Brown MR, Hakim FT, Bare CV, Leitman SF, Read EJ, Carter CS, Wexler LH, Gress RE. Prolonged CD4 depletion after sequential autologous peripheral blood progenitor cell infusions in children and young adults. Blood. 2000;96:754–762. - PubMed
-
- Connors M, Kovacs JA, Krevat S, Gea-Banacloche JC, Sneller MC, Flanigan M, Metcalf JA, Walker RE, Falloon J, Baseler M, Feuerstein I, Masur H, Lane HC. HIV infection induces changes in CD4+ T-cell phenotype and depletions within the CD4+ T-cell repertoire that are not immediately restored by antiviral or immune-based therapies. Nat Med. 1997;3:533–540. doi: 10.1038/nm0597-533. - DOI - PubMed
-
- Lum LG. The kinetics of immune reconstitution after human marrow transplantation. Blood. 1987;69:369–380. - PubMed
-
- Ochs L, Shu XO, Miller J, Enright H, Wagner J, Filipovich A, Miller W, Weisdorf D. Late infections after allogeneic bone marrow transplantations: comparison of incidence in related and unrelated donor transplant recipients. Blood. 1995;86:3979–3986. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
